 2014-02-13
2014-02-13 4429
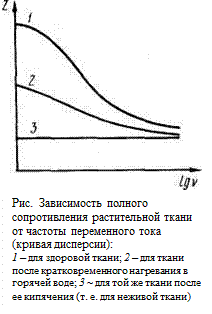
4429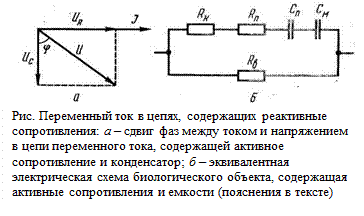 Как известно, при прохождении переменного тока в цепи, состоящей из активного, емкостного и индуктивного сопротивлений, сила тока в цепи I и напряжение U связаны обобщенным законом Ома: I = U/z, где z – полное сопротивление, т. е. величина, учитывающая все виды сопротивлений цепи переменному току. В биообъектах индуктивность практически отсутствует, и величина полного сопротивления определяется только активным сопротивлением и емкостью, которые образуют более или менее сложные, так называемые эквивалентные цепи. В случае если активное сопротивление R и емкость С соединены последовательно, то величину полного сопротивления вычисляют по формуле:
Как известно, при прохождении переменного тока в цепи, состоящей из активного, емкостного и индуктивного сопротивлений, сила тока в цепи I и напряжение U связаны обобщенным законом Ома: I = U/z, где z – полное сопротивление, т. е. величина, учитывающая все виды сопротивлений цепи переменному току. В биообъектах индуктивность практически отсутствует, и величина полного сопротивления определяется только активным сопротивлением и емкостью, которые образуют более или менее сложные, так называемые эквивалентные цепи. В случае если активное сопротивление R и емкость С соединены последовательно, то величину полного сопротивления вычисляют по формуле:
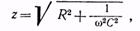
где w = 2 pv – циклическая частота переменного тока, а R с = 1/ wС – емкостное сопротивление.
В цепях переменного тока, в которых существуют реактивные, т. е. емкостные и индуктивные сопротивления, сила тока и напряжение сдвинуты по фазе на некоторый угол ф. При последовательном соединении активного и емкостного сопротивлений (рис. а) токи IR и I с, проходящие через оба сопротивления, равны и тангенс угла сдвига фаз, равный отношению падений напряжений на реактивном и на активном сопротивлениях
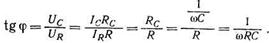
Как было показано, емкость биологических объектов определяется статической емкостью клеточных мембран С ми поляризационной емкостью С п, которые можно считать соединенными последовательно. При последовательном соединении результирующая емкость может быть найдена по формуле
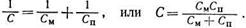
Поляризационная емкость зависит от времени, в течение которого действует электрическое поле. При значительном времени, а следовательно, при малой частоте переменного поля величина поляризационной емкости может быть значительной и даже превышать статическую емкость. При высоких частотах переменного поля величина поляризационной емкости снижается и при частоте более 10 кГц она становится на несколько порядков меньше величины статической емкости, т. е. С п <<С М. Из последней формулы видно, что при этом С» Сп, т. е. результирующая емкость биообъекта почти полностью определяется меньшей по величине поляризационной емкостью.
Эквивалентная электрическая схема биологического объекта показана на (рис. 6). На этой схеме R П–активное сопротивление, обусловленное поляризационными явлениями на клеточных мембранах. Величина поляризационного сопротивления, так же как и поляризационная емкость, зависит от величины результирующих ионных потоков через мембрану, и следовательно, от частоты переменного тока и от проницаемости клеточных мембран. Последовательно с поляризационной емкостью и сопротивлением включено активное сопротивление клеточной цитоплазмы R K. Помимо клетки, ток проходит и по внеклеточной жидкости, и поэтому все внутриклеточные сопротивления шунтируются сопротивлением внеклеточной среды R B. Кроме R Пи С п, все остальные элементы эквивалентной схемы от частоты не зависят.
Рассмотрим, как будет меняться полное сопротивление биообъекта при изменении частоты переменного тока. При низких частотах поляризационные эффекты почти так же велики, как при постоянном токе; соответственно велики R пи С п. При больших межклеточных промежутках их сопротивление мало и практически весь ток пойдет по шунтирующему сопротивлению R B. Если же сечение межклеточных промежутков невелико, то сопротивление R Bвозрастает и измеряемое полное сопротивление будет в основном определяться величиной R п. Поскольку R пзависит от проницаемости мембран, то величина сопротивления объекта, измеренная на низких частотах, в ряде случаев может служить мерой проницаемости клеточных мембран. При возрастании частоты переменного тока поляризационные эффекты уменьшаются, соответственно уменьшаются величины и С п - При частотах порядка нескольких МГц поляризация мембран почти исчезает и величинами R пи С п при расчете полного сопротивления можно пренебречь. Таким образом, полное сопротивление, измеренное на высоких частотах, будет равно
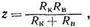
так как оно будет определяться параллельно соединенными сопротивлениями электролитов внутри и вне клетки. Отсюда следует, что высокочастотное сопротивление биообъектов может служить мерой концентрации в них свободных ионов.
При дальнейшем возрастании частоты (до 1 ГГц и выше) уменьшается поляризация, связанная с ориентацией дипольных молекул воды, что ведет к падению величины диэлектрической проницаемости, которая при измерениях на сверхвысоких частотах значительно меньше, чем на низких частотах, поскольку на высоких частотах она определяется только электронной и ионной поляризациями.
Таким образом, электросопротивление (или электропроводность) тесно связано как со свойствами клеточных мембран, так и со свойствами клеточных и межклеточных жидкостей. Как было показано выше, измерение удельного сопротивления у живых объектов представляет большие трудности, однако для диагностических целей это и не требуется. Получать сведения о явлениях, происходящих в тканях, можно, наблюдая за относительным изменением их электросопротивления между электродами любой формы, что широко применяют в медицине, например, для диагностики воспалительных процессов. В начальной стадии воспаления структура клеток заметно не меняется и, следовательно, не меняется их электроемкость, однако происходит набухание клеток и тканей, уменьшается сечение межклеточных промежутков, что ведет к возрастанию активного сопротивления. Как говорилось ранее, при измерениях на низких частотах основное сопротивление ткани определяется именно сопротивлением межклеточных промежутков. Поэтому, измеряя сопротивление на низких частотах, по значительному возрастанию сопротивления можно судить о начале воспалительного процесса. При дальнейшем развитии воспаления химический состав и структура клеток изменяются, увеличивается проницаемость мембран для ионов, что ведет к уменьшению электроемкости и сопротивления клеток и в конечном счете к уменьшению их полного сопротивления. Таким образом, сильное уменьшение электросопротивления ткани на низких частотах может свидетельствовать о развитом воспалительном процессе.
 Весьма важные сведения о состоянии биологических объектов дают дисперсионные кривые, т. е. графики зависимости полного сопротивления ткани от частоты переменного тока. Такая зависимость присуща только живым тканям; при пропускании переменного тока через обычные растворы электролитов дисперсия электросопротивления не наблюдается. Причина дисперсии заключается в том, что на величину электросопротивления при постоянном токе или при низких частотах значительное влияние оказывает макроструктурная поляризация. Поскольку при увеличении частоты переменного тока уменьшаются пограничные поляризационные эффекты, это ведет к уменьшению полного сопротивления ткани, и дисперсионная кривая обладает значительной крутизной (рис. кривая 1). При повреждении ткани возрастает проницаемость клеточных мембран, что приводит к уменьшению R пи С пи, следовательно, к уменьшению полного сопротивления на низких частотах. Поэтому кривая 2на рисунке обладает меньшей крутизной. При отмирании ткани поляризация на границах раздела практически исчезает и зависимость электросопротивления от частоты отсутствует, так как остается лишь активное сопротивление (рис. кривая 3). Таким образом, по крутизне дисперсионной кривой можно судить о жизнеспособности той или иной ткани, что имеет большое значение, когда эта ткань предназначена для трансплантации.
Весьма важные сведения о состоянии биологических объектов дают дисперсионные кривые, т. е. графики зависимости полного сопротивления ткани от частоты переменного тока. Такая зависимость присуща только живым тканям; при пропускании переменного тока через обычные растворы электролитов дисперсия электросопротивления не наблюдается. Причина дисперсии заключается в том, что на величину электросопротивления при постоянном токе или при низких частотах значительное влияние оказывает макроструктурная поляризация. Поскольку при увеличении частоты переменного тока уменьшаются пограничные поляризационные эффекты, это ведет к уменьшению полного сопротивления ткани, и дисперсионная кривая обладает значительной крутизной (рис. кривая 1). При повреждении ткани возрастает проницаемость клеточных мембран, что приводит к уменьшению R пи С пи, следовательно, к уменьшению полного сопротивления на низких частотах. Поэтому кривая 2на рисунке обладает меньшей крутизной. При отмирании ткани поляризация на границах раздела практически исчезает и зависимость электросопротивления от частоты отсутствует, так как остается лишь активное сопротивление (рис. кривая 3). Таким образом, по крутизне дисперсионной кривой можно судить о жизнеспособности той или иной ткани, что имеет большое значение, когда эта ткань предназначена для трансплантации.
Частотные зависимости электросопротивления сходны для многих тканей, однако величины полного сопротивления разных тканей различны. Так, например, костная ткань, содержащая большое количество кристаллов фосфата кальция, обладает большим удельным сопротивлением, чем мягкие ткани. Зона дисперсии электросопротивления обычно находится в пределах от 0,1 кГц до 100 МГц, однако у некоторых тканей минимальное сопротивление достигается еще на больших частотах (например, у нервной ткани около 1 ГГц). Крутизну кривой дисперсии принято характеризовать отношением величин полных сопротивлений ткани, измеренных на низких и высоких частотах в одинаковых условиях. Наиболее удобны частоты 104 Гц, так как при этой частоте обычно наблюдается перелом кривой дисперсии, и 106 Гц, так как для большинства тканей при этом достигается минимальное сопротивление. Как показывает опыт, это отношение, называемое коэффициентом поляризации К, для данной ткани в норме есть величина практически постоянная
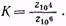
Величина К зависит от интенсивности обмена веществ в ткани, а также для тканей одинаковых органов от степени эволюционного развития животного. Например, для печени лягушки К = 2–3, а для печени млекопитающих К = 9–10. После отмирания ткани величина К приближается к единице.
Имеет также диагностическое значение измерение угла сдвига фаз между током и напряжением. Этот угол для биообъектов довольно большой, что указывает на значительную долю емкостного сопротивления. Так, измерения tg j для различных объектов показали, что на частоте 1 МГц у нерва лягушки j = 64°, у мышцы кролика j = 65°, у кожи человека j = 55° и т.п. Для других частот сдвиг фаз будет иным, хотя он и не столь сильно зависит от частоты, как величина полного сопротивления. При патологии величина j изменяется, что также может служить для целей диагностики.
Ниже приведены значения углов сдвига фаз, полученные при частоте 1 кГц для разных биологических объектов.
| Биологический объект | Угол сдвига фаз, град |
| Кожа человека, лягушки | |
| Нерв лягушки | |
| Мышцы кролика | |
| Десна | |
| Эмаль зуба |
Полное сопротивление тканей живого организма в значительной степени зависит от состояния кровообращения, от степени наполнения кровеносных сосудов, проходящих в этих тканях. Кровь имеет меньшее удельное сопротивление, чем стенки сосудов или клетки, и поэтому при наполнении ткани кровью во время систолы полное сопротивление ткани уменьшается, а при диастоле увеличивается. Исследование периферического кровообращения путем измерения электросопротивления тканей называют реографией. Для реографии применяют переменный ток с частотой 20–30 кГц и измеряют полное сопротивление определенного участка тканей в течение цикла сердечной деятельности. Методом реографии исследуют сосуды головного мозга (реоэнцефалография), легких (реопульмонография), глаз (реоофтальмография), конечностей (реовазография). Электроды при этом накладывают на соответствующий орган, смазав предварительно специальной пастой для улучшения контакта с кожей. Относительное изменение объема ткани при кровенаполнении D V/V равно относительному изменению величины полного сопротивления D z/z:
 .
.
Знак минус означает, что при увеличении кровенаполнения во время систолы сопротивление ткани уменьшается.
Приборы для получения реограмм позволяют вычерчивать пульсовую волну сопротивления, причем каждый орган в отдельности в норме дает вполне определенное значение изменения сопротивления. Так, амплитудные значения D z для человека: плечо – 0,05 Ом, кисть руки – 0,1, бедро – 0,035 Ом и т. п. Исследование реограмм применяют в диагностике заболеваний периферических кровеносных сосудов, сопровождающихся изменением их эластичности, сужением артерий и прочим, а также при исследовании механизма экстремальных воздействий на организм, действия лекарственных веществ и т. п.
Дисперсия импеданса. В реальных биологических тканях имеются отличия в частотной зависимости импеданса.
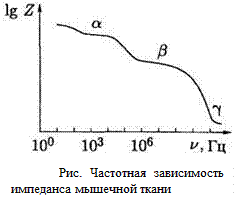 Общих ход зависимости свидетельствует о плавном уменьшении импеданса с увеличением частоты. При этом четко проявляются три интервала частот, в которых величина Z медленнее меняется с частотой по сравнению с общим ходом кривой. Они названы, соответственно, областями a, b, и g -дисперсии.
Общих ход зависимости свидетельствует о плавном уменьшении импеданса с увеличением частоты. При этом четко проявляются три интервала частот, в которых величина Z медленнее меняется с частотой по сравнению с общим ходом кривой. Они названы, соответственно, областями a, b, и g -дисперсии.
При воздействии переменным (гармоническим) электрическим полем проявляется частотная зависимость диэлектрической проницаемости: е = /(^). Изменение диэлектрической проницаемости с изменением частоты электрического поля приводит к изменению электроемкости, а следовательно, к изменению импеданса (теоретическое доказательство этого здесь не приводится)
На рис. показана частотная зависимость диэлектрической проницаемости, в которой также выделяются области a, b, и g -дисперсии.
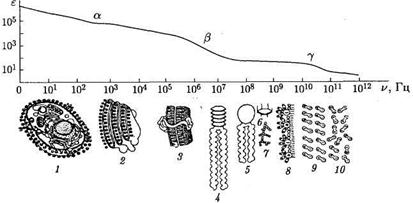
Рис. Частотная зависимость диэлектрической проницаемости
Наличие областей дисперсии указывает на различные механизмы поляризации тканей в разных частотных диапазонах действующего внешнего электромагнитного поля. Каждый из механизмов поляризации характеризуется своей частотой, вблизи которой запаздывание смещения (поворота) различных элементов биологических тканей относительно изменения действующего переменного электромагнитного поля минимально.
Механизмы возникновения областей дисперсии следующие:
a-дисперсия обусловлена поляризацией целых клеток (1, 2)в результате диффузии ионов, что требует относительно большого времени, поэтому данный механизм проявляется при действии электрического поля низкой частоты (0,1-10 кГц); в этой области емкостное сопротивление мембран очень большое и преобладают токи, протекающие через растворы электролитов, окружающие фрагменты мембран.
b-дисперсия обусловлена структурной поляризацией клеточных мембран (3),в которой участвуют белковые макромолекулы (4), а на их верхней границе – глобулярные водорастворимые белки (5), фосфолипиды (6, 7)и мельчайшие субклеточные структуры (8);данный механизм проявляется при действии переменного электрического поля с частотой 1-10 МГц;
g-ducnepcuя обусловлена процессами ориентационной поляризации молекул (9, 10) свободной и связанной воды (частота около 20 ГГц), а также низкомолекулярных веществ типа сахаров и аминокислот (частотный интервал 1-10 ГГц).
По частотной зависимости импеданса можно оценивать жизнеспособность ткани, что важно при пересадке органов. Различия в частотных зависимостях импеданса имеют место при сравнении здоровой и больной ткани.








