 2015-01-21
2015-01-21 877
877Явление коацервации состоит в том, что в некоторых условиях (например, в присутствии электролитов) высокомолекулярные вещества отделяются от раствора, но не в форме осадка, а в виде более кон центрирован но го раствора — коацервата. При встряхивании коацерват распадается на отдельные мелкие капельки. В воде такие капли покрываются стабилизирующей их гидратной оболочкой (оболочкой из молекул воды) — рис. 2.4.1.4.
Коацерватные капли обладают некоторым подобием обмена веществ: иод воздействием чисто физико-химических сил они могут избирательно впитывать из раствора некоторые вещества и выделять в окружающую среду продукты их распада. За счет избирательного концентрирования веществ из окружающей среды они могут расти, а но достижении определенного размера начинают «размножаться», отпочковывая маленькие капельки, которые, в свою очередь, могут расти и «почковаться».
Возникшие в результате концентрирования белковых растворов коацерватные капли в процессе перемешивания под действием волн и ветра могут покрываться оболочкой из липи- дов: одинарной, напоминающей мицеллы мыла (при однократном отрыве капли от поверхности воды, покрытой липидным слоем), либо двойной, напоминающей клеточную мембрану (при повторном падении капли, покрытой однослойной липидной мембраной, на липидную пленку, покрывающую поверхность водоема — рис. 2.4.1.4).
Процессы возникновения коацерватных капель, их роста и «почкования», а также «одевания» их мембраной из двойного липидного слоя легко моделируются в лабораторных условиях.
Для коацерватных капель также существует процесс «естественного отбора», при котором в растворе сохраняются наиболее стабильные капли.
Несмотря на внешнее сходство коацерватных капель с живыми клетками, у коацерватных капель отсутствует главный признак живого — способность к точному самовоспроизведению, самокопированию. Очевидно, предшественниками живых клеток явились такие коацерватные капли, в состав которых вошли комплексы молекул-репликаторов (РНК или ДНК) и кодируемых ими белков. Возможно, комплексы РНК-белок длительное время существовали вне коацерватных капель в виде так называемого «свободноживущего гена», а возможно, их формирование проходило непосредственно внутри некоторых коацерватных капель.
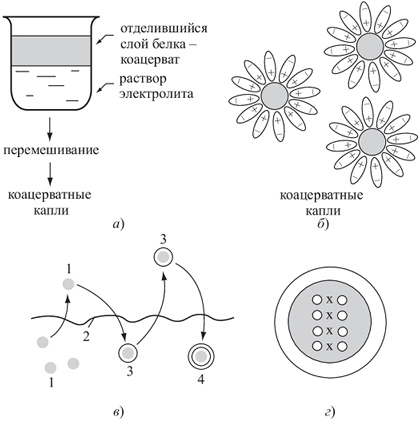
Рис 2.4.1.4.
Возможный путь перехода от коацерватных капель к примитивным клешам:
А) образование коацсрвата; 6) стабилизация коацерватных капель в водном растворе; в) — формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану: 1 — коацерватная капля; 2 — мономолекулярный слой липида на поверхности водоема; 3 — формирование вокруг капли одинарного липидного слоя; 4 — формирование вокруг капли двойного липидного слоя, похожего на клеточную мембрану; г) — коацерватная капля, окруженная двойным липидным слоем, с вошедшим в ее состав белково-нуклеотидным комплексом — прообраз первой живой клетки
Исключительно сложный, не до конца понятный современной науке процесс возникновения жизни на Земле прошел с исторической точки зрения чрезвычайно быстро. Уже 3,5 млрд лет т.н. химическая эволюция завершилась появлением первых живых клеток и началась биологическая эволюция.
Начальные этапы биологического обмена. Синтетические процессы, протекающие в абиогенной среде, могли быть усилены в случае их пространственной локализации. Так, в опытах с коацерватами – каплями, обогащенными веществом (от лат. coacervatio – накопление), показано, что при помещении их в раствор глюкозо-1-фосфата (Г-1-Ф) они способны интенсивно синтезировать крахмал (если предварительно в коацерваты была введена фосфорилаза). После одновременного введения фосфорилазы и β-амилазы в коацерваты и помещения последних в раствор Г-1-Ф вначале синтезировался крахмал, а затем мальтоза.
Полагают, что в начальных этапах предбиологической эволюции коацерваты, в которых возникла концентрация определенного набора металлов и простых органических веществ, стали основой для перехода к биогенным синтезам. Эти синтезы усиливались и принимали более направленный характер с появлением в коацерватах и АТФ, синтезированной первоначально абиогенным путем. Даже первые живые организмы могли получать АТФ из окружающей среды.
С другой стороны, каталитическая активность коацерватов могла быть усилена и при возникновении комплексов между металлами и органическими соединениями. Так, с внедрением в порфириновое кольцо ионов железа его каталитическая активность возрастает в 100 раз по сравнению с ионами железа в растворе. В результате такого сочетания, вероятно, появились коферменты в коацерватах, а затем и субстрат-специфические ферменты, способствовавшие биогенным синтезам. При возникновении ферментов исходно могли быть использованы простые пептиды в комплексе с ионами металлов.
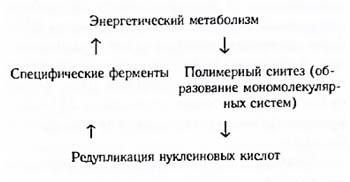 До возникновения истинно живого существа способность к самовоспроизведению не обязательно могла быть связана с точной редупликацией нуклеиновых кислот, так как она осуществлялась очень медленно. Самоудвоение их могло бы быть результатом постоянства соотношения скоростей разных реакций обмена веществ, идущих с участием коферментов – катализаторов небелковой природы. В последующем, видимо, шел предбиологический отбор коацерватов (А.И. Опарин) и по способности накопления специальных белковоподобных полимеров, ответственных за ускорение химических реакций. Результаты такого отбора могли фиксироваться в строении нуклеиновых кислот путем преимущественного размножения систем, в которых удвоение нуклеиновых кислот осуществлялось с участием ферментов. Все эти изменения привели к возникновению циклического обмена веществ, характерного для живых существ:
До возникновения истинно живого существа способность к самовоспроизведению не обязательно могла быть связана с точной редупликацией нуклеиновых кислот, так как она осуществлялась очень медленно. Самоудвоение их могло бы быть результатом постоянства соотношения скоростей разных реакций обмена веществ, идущих с участием коферментов – катализаторов небелковой природы. В последующем, видимо, шел предбиологический отбор коацерватов (А.И. Опарин) и по способности накопления специальных белковоподобных полимеров, ответственных за ускорение химических реакций. Результаты такого отбора могли фиксироваться в строении нуклеиновых кислот путем преимущественного размножения систем, в которых удвоение нуклеиновых кислот осуществлялось с участием ферментов. Все эти изменения привели к возникновению циклического обмена веществ, характерного для живых существ:
В системе коацерватов не исключен и отбор самих нуклеиновых кислот по наиболее удачному сочетанию смысловой последовательности нуклеотидов, т.е. отбор по генам. Первый отбор самореплицирующихся молекул РНК должен был идти по скорости и точности репликации – для построения копий нужны были запасы нуклеотидов (аналогия с конкуренцией организмов за пищевые ресурсы). Системы с удачно работающей (смысловой) последовательностью нуклеотидов в нуклеиновой кислоте вправе называться живыми.
Другие гипотезы возникновения жизни на Земле.Первые водоемы появились 3,8–4 млрд лет назад, и тогда же в них отложились первые осадочные породы (глинистые минералы). Гипотеза Дж. Бернала говорит об огромной роли глинистых отложений в концентрации органики абиогенного происхождения, а также о значении межзвездного вещества (цианистый водород, оксид углерода, формальдегид и др.), доставляемого на поверхность Земли кометами и метеоритами.
Земля, проходя через пылевое облако в течение 105–106 лет, могла получить вместе с космической пылью 108–1010 т органического материала, что превосходит в количественном отношении современную биомассу нашей планеты. Органическое вещество Земли пополнялось и за счет извержения вулканов. Так, подсчитано, что одно извержение вулкана выбрасывает на Землю до 1000 т органического вещества. Благодаря оседанию на поверхности ультрамикроскопических пылинок разные компоненты космической или вулканической органики могли вступить между собой в реакции, образуя своеобразную оболочку из аминокислот, мочевины и других веществ, используя энергию УФ-лучей. Подобные процессы создавали необходимую концентрацию веществ пространственно и могли способствовать быстрому возникновению живых существ в первичном океане.
Теоретически можно представить и другие пути возникновения жизни. Например, при попадании воды с растворенными в ней аминокислотами на горячий субстрат (мелкие водоемы в застывающей лаве) мог осуществиться процесс полимеризации с образованием протеиноидных микросфер (эксперименты К. Фолсома и Р. Фокса). Сами аминокислоты могли бы выступать в роли катализаторов собственных реакций при образовании полипептидов. Ту же каталитическую роль в возникновении протобионтов могли выполнять молекулы РНК и полипептиды. Способствовать этим процессам могло «самоконструирование» макромолекул (М. Эйген) при объединении их в более сложные комплексы.
В протобионтах каждый блок или молекула РНК должны были заново синтезироваться из диспергированных в воде молекул. Эра биологической эволюции началась тогда, когда каким-то образом возникли гены.
Хиральная чистота живого.В вопросе о происхождении жизни одним из загадочных остается факт наличия абсолютной хиральной чистоты (от греч. cheir – рука): у живых существ – содержание в молекулах белков только «левых» аминокислот, а в нуклеиновых кислотах – «правых» сахаров. Подобное явление могло возникнуть только вследствие утраты предбиологической средой первичной зеркальной симметрии (равное содержание правых и левых изомеров аминокислот и сахаров). Неживой природе присуща тенденция установления зеркальной симметрии (рацемации).
Опыты последних лет показали, что только в хирально чистых растворах практически могли возникнуть биологически значимое удлинение цепочки полинуклеотидов и процесс саморепликации. Рацемический полинуклеотид не в состоянии реплицироваться, так как его основания направлены в разные стороны и у него нет спиральной организации. Живые системы организованы так, что тРНК из правых сахаров присоединяют к себе только левые аминокислоты. Поэтому возникновение жизни, по-видимому, исключалось до разрушения зеркальной симметрии предбиологической среды (В. Гольданский, Л. Морозов) и появления самореплицирующихся систем. Все живые организмы поддерживают свою хиральную чистоту, и эволюция не снабдила их средствами для обитания в рацемической среде.
4) Кризис классического дарвинизма. Несмотря на признание учения Ч. Дарвина большинством биологов, многие ведущие биологи либо не приняли (Р. Оуэн, К. Бэр), либо принимали учение об эволюции путем естественного отбора с оговорками, либо выдвигали серьезные возражения (Л. Агассиц, Л. Келликер, К. Негели и др.). Этому в немалой степени способствовало то, что взгляды Ч. Дарвина на всемогущество отбора и особенности возникновения новых видов встретились с рядом трудностей (например, в природе редко находились переходные формы, присутствие которых постулировал Дарвин), возникновение органов очень сложного строения и сложных форм поведения также, на первый взгляд, было трудно объяснить посредством аккумуляции мелких изменений (недостаточным казался принцип отбора и для объяснения первых стадий развития полезных уклонений, неясным оставалось возникновение общего плана строения в крупных группах организмов и т.п.).
Критика дарвинизма, особенно усилилась в период возникновения генетики. Этот период условно можно назвать периодом отрицания дарвинизма. В это время (конец XIX– начало XX в.) не только в биологии, но и в других областях естествознания нападки на материалистические взгляды усилились. Судьбы эволюционного учения и теории естественного отбора разошлись. Эволюционное учение распространялось все шире и шире, а теория естественного отбора – центральное звено всей эволюционной теории – стала подвергаться все более жесткой критике. Причин тому было несколько, но основная – отсутствие достаточно строгих научных представлений о двух важнейших явлениях жизни и общих характеристиках живого на Земле – наследственности и изменчивости.
Вскоре после выхода «Происхождения видов...» Ф. Дженкин выдвинул серьезное возражение против предполагаемой Ч. Дарвином возможности действия отбора как эволюционного фактора в природе. Ход его рассуждений был следующим. Возникшее случайное наследственное изменение, которое должно быть поддержано отбором, всегда единично. Вероятность встречи двух особей с одинаковыми наследственными изменениями и оставлением ими потомства чрезвычайно мала. Поэтому если один из родителей имеет признак А, то у его детей количественное выражение признака будет А/2, у внуков – А/4, у правнуков – А/8 и т.д., т.е. произойдет «растворение признака в скрещивании».
Для опровержения возражения Дженкина необходимо было знание генетики, а генетика как наука возникла лишь в 1900 г., в год переоткрытия замечательных работ Г. Менделя. Правда, и ранее были известны многочисленные факты, свидетельствовавшие против «растворения признаков в скрещивании». Например, характерная горбинка на носу у представителей королевского дона Бурбонов сохранилась даже у восьмого по счету поколения. По расчетам Ф. Дженкина, признак должен был уменьшиться в 128 раз. Четырнадцатый по счету поколений герцог Шрюсбери имел, как и основоположник рода (500 лет назад), сросшиеся первые и вторые фаланги на пальцах рук. Но эти факты (как и аналогичные известные тогда факты из области селекции) оставались вне внимания исследователей – эволюционистов.
Существовал и другой фронт исследований, прямо или косвенно противопоставляемый дарвинизму в конце XIX – начале XX в. В работе В. Иоганнсена (1903) была показана неэффективность отбора в чистых линиях (в потомстве одной самооплодотворяющейся особи, гомозиготном по большинству признаков). И хотя такие гомозиготные группы особей в природе не встречаются, веру в могущество отбора эти эксперименты подрывали. Крайне неудачная «временная гипотеза пангенезиса», предложенная Дарвином, расходилась с наблюдаемыми фактами наследственной передачи признаков в экспериментах. Это было, по-видимому, одной из причин выдвижения Г. де Фризом (1889) гипотезы эволюции, согласно которой новые виды возникают сразу, скачкообразно, посредством появления отдельных крупных изменений наследственности (мутации) без ведущего участия естественного отбора.
Причиной указанных трудностей дарвинизма, как уже отмечалось, было отсутствие достаточно строгих доказательств наследственной изменчивости, теории отбора «не хватало» генетической основы. Такие достижения генетики, как мутационная теория, учение о чистых линиях и принцип корпускулярной наследственности, использовались для обоснования антидарвиновских взглядов. Эти данные генетики на самом деле укрепляли учение Ч. Дарвина, внося ясность в понимание механизма действия естественного отбора.
Генетики того периода вместо расплывчатых представлений Ч. Дарвина и его сторонников о наследственнocти пытались предложить твердые законы и основанные на экспериментах гипотезы. Этот подход просмотрели сторонники взглядов Ч. Дарвина, которые, в свою очередь, предпринимали безуспешные попытки опровергнуть ранних генетикой. Генетики, со своей стороны, увлеченные успехами, не заметили положительное в учении Ч. Дарвина. Это и было причиной конфликта. Знаменательным событием в истории развития эволюционного учения стал 1926 год – год появления работы С.С. Четверикова «О некоторых моментах эволюционного процесса с точки зрения современной генетики», давшей начало синтезу генетики и классического дарвинизма.
Период синтеза генетики и классического дарвинизма.Непростым был путь к синтезу. Первым шагом на этом пути может считаться вскрытие закономерностей распределения хромосом при клеточном делении. Основываясь на этих фактах, А. Вейсман (1834–1914) формулирует основные положения хромосомной теории наследственности и впервые выдвигает принцип невозможности передачи по наследству «благоприобретенных» признаков.
До сих пор иногда встречаются исследователи, признающие принцип наследования «благоприобретенных» признаков («адекватная изменчивость», «соматическая индукция» и т.п.). Однако из признания факта влияния внешней среды на наследственность (никогда не оспариваемого генетикой) не вытекает возможность адекватной изменчивости. По-прежнему справедливо прекрасное сравнение Ч. Дарвина: «...мы ясно видим, что природа условий имеет в определении каждого данного изменения подчиненное значение по сравнению с природой самого организма; быть может, она имеет не большее значение, чем имеет природа той искры, которая воспламеняет массу горючего материала, в определении свойства (вспыхивающего) пламени».
Упоминавшаяся мутационная гипотеза эволюции Г. де Фриза сыграла важную положительную роль при синтезе генетики и дарвинизма, ускорив накопление точных данных по наследственной («неопределенной», по Дарвину) изменчивости в живой природе. После обнаружения у дрозофилы групп сцепления генов по числу имеющихся хромосом окончательно оформляется хромосомная теории наследственности (Т.Г. Морган, А. Стертевант и др.).
Переход к популяционному мышлению.В потоке разнообразных генетических исследований формируется знаменитый «принцип Харди» (1908): без внешнего давления каких-либо факторов частоты генов в бесконечно большой панмиктической популяции стабилизируются уже после одной смены поколений. Другими словами, при отсутствии внешних давлений частоты генов в популяции должны быть постоянными.
Исходя из принципа Харди, С.С. Четвериков (1882–1959) показал, что в результате постоянно протекающего мутационного процесса во всех популяциях создается и существует наследственная гетерогенность – различные мутации и комбинации, представляющие генетическую основу эволюционного процесса. Из расчетов С.С. Четверикова следовало, что во всех без исключения популяциях должны присутствовать самые различные мутации. В ходе «переработки» этих мутаций под действием естественного отбора и осуществляется процесс эволюции. Экспериментальные проверки полностью подтвердили вывод С.С. Четверикова о насыщенности природных популяций разными мутациями.
Проведенные в 1928–1930 гг. исследования (Р.А. Фишер, Н.П. Дубинин и Д.Д. Ромашов, С. Раит и др.) показали, что в эволюции большую роль играет не только появление новых мутаций, но и изменение частоты встречаемости существующих аллелей (гена) благодаря случайным процессам – колебания численности популяции, утраты генов и т.д.
Генетика позволила проанализировать основные моменты протекания эволюционного процесса от появления нового признака в популяции до возникновения нового вида. На внутривидовом (микроэволюционном) уровне при изучении эволюции оказалось возможным применить точные экспериментальные подходы, которые помогли выяснить роль отдельных эволюционных факторов, сформулировать представления об элементарной эволюционной единице (популяция), элементарном эволюционном материале и явлении. Все это привело к созданию Ф.Г. Добржанским и Н.В. Тимофеевым-Ресовским в 1937–1939 гг. учения о микроэволюции – одного из главных разделов современной теории эволюции.
Неодарвинизм (вэйсманизм) — эволюционная концепция, созданная А. Вейсманом на раннем этапе развития генетики (в кон. XIX — нач. XX вв.). Неодарвинизмом иногда называют современный дарвинизм, представляющий собой синтез генетики и классического дарвинизма, что неправильно с исторической точки зрения[1]








