 2015-02-14
2015-02-14 746
746Потребность организма в энергии характеризуется таким уровнем ее потребления с пищей, при котором на фоне неизменной массы тела, физической активности и соответствующих скоростях роста и обновления организма достигается энергетический баланс поступления и расхода энергии. Живые организмы получают энергию в виде потенциальной энергии питательных веществ. Эта энергия аккумулирована в химических связях молекул жиров, белков и углеводов, которые в процессе катаболизма преобразуются в конечные продукты обмена с более низким содержанием энергии. Высвобождающаяся в процессе биологического окисления энергия используется, прежде всего, для синтеза АТФ, которая как универсальный источник энергии, необходима в организме для последующего осуществления механической работы, химического синтеза и обновления структур, транспорта веществ, осмотической и электрической работы. (см.рис.10.2). Количество синтезированных молей АТФ на моль окисленного субстрата зависит от его вида и от величины коэффициента фосфорилирования. Этот коэффициент, обозначаемый как Р/ О, равен количеству синтезированных молекул АТФ в расчете на один атом кислорода, потребленный при окислении восстановленных органических соединений в процессе дыхания. При переносе каждой пары электронов по дыхательной цепи от НАД • Н до О2 величина Р/О = 2. Для субстратов, окисляемых ФАД • Н2 — зависимыми ферментами Р/О = 1,3. Эти соотношения Р/О отражают энергетические затраты клетки на синтез АТФ в митохондриях и транспорт макроэрга против химического градиента из митохондрий к местам потребления.
Таким образом, одна часть аккумулированной в химических связях молекул жиров, белков и углеводов энергии в процессе биологического окисления используется для синтеза АТФ, другая часть этой энергии превращается в теплоту. Эта теплота, выделяющаяся сразу же в процессе биологического окисления питательных веществ, получила название первичной. Какая часть энергии будет использована на синтез АТФ и будет вновь аккумулирована в ее химических макроэргических связях зависит от величины Р/О и эффективности сопряжения в митохондриях процессов дыхания и фосфорилирова-
460



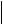 ния. Разобщение дыхания и фосфорилирования под действием гормонов щитовидной железы, ненасыщенных жирных кислот, липо-протеидов низкой плотности, динитрофенола ведет к уменьшению коэффициента Р/О, превращению в первичную теплоту большей, чем в условиях нормального сопряжения дыхания и фосфорилирования части энергии химических связей окисляемого вещества. При этом снижается коэффициент полезного действия синтеза АТФ, количество синтезированных молекул АТФ уменьшается.
ния. Разобщение дыхания и фосфорилирования под действием гормонов щитовидной железы, ненасыщенных жирных кислот, липо-протеидов низкой плотности, динитрофенола ведет к уменьшению коэффициента Р/О, превращению в первичную теплоту большей, чем в условиях нормального сопряжения дыхания и фосфорилирования части энергии химических связей окисляемого вещества. При этом снижается коэффициент полезного действия синтеза АТФ, количество синтезированных молекул АТФ уменьшается.
Наиболее ярким примером преимущественного превращения энергии химических связей окисляемого жира в первичную теплоту, а не на синтез АТФ и получения быстрого калоригенного эффекта является разобщающее действие полипетида молекулярной массой 32000 в бурой жировой ткани.
В нормальных условиях при полном окислении 1 г смеси углеводов пищи выделяется 4 ккал тепла. В процессе окисления в организме 1 г углеводов синтезируется 0,13 моля АТФ. Если считать, что энергия пирофосфатной связи в АТФ равна 7 ккал/моль, то при окислении 1 г углеводов лишь 0,91 (0.13 х 7) ккал энергии будет запасено в организме в синтезированной АТФ. Остальные 3,09 ккал будут рассеяны в виде тепла (первичная теплота). Отсюда можно рассчитать коэффициент полезного действия синтеза АТФ и аккумулирования в ней энергии химических связей глюкозы.
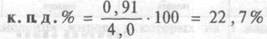
Из приведенного расчета видно, что только 22,7% энергии химических связей глюкозы в процессе ее биологического окисления используется на синтез АТФ и вновь запасается в виде химической макроэргической связи, 77,3% энергии химических связей глюкозы превращается в первичную теплоту и рассеивается в тканях.
Процессы использования энергии в тепло можно представить на схеме (рис. 10.2). Из нее видно, что аккумулированная в АТФ энергия в последующем используется для осуществления в организме химических, транспортных, электрических процессов, производства механической работы и в конечном итоге тоже превращается в теплоту, получившую название вторичной.
В названиях первичная и вторичная теплота отражено представление о двухступенчатости полного превращения всей энергии химических связей питательных веществ в тепло (первая ступень — образование первичной теплоты в процессе биологического окисления, вторая ступень — образование вторичной теплоты в процессе затраты энергии макроэргов на производство различных видов работы). Таким образом, если измерить все количество тепла, образовавшегося в организме за час или сутки, то это тепло станет мерой суммарной энергии химических связей питательных веществ, подвергшихся за время измерения биологическому окислению. По количеству образовавшегося в организме тепла можно судить о величине энергетических затрат, произведенных на осуществление процессов жизнедеятельности.
461






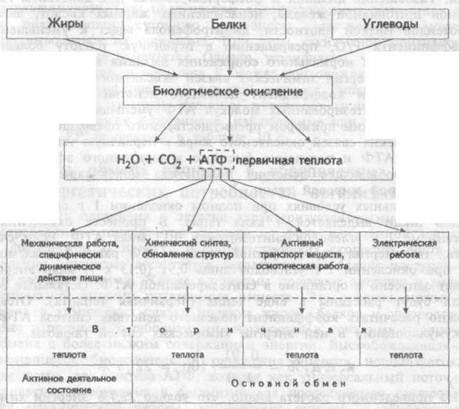
Рис.10.2. Схематическое представление взаимосвязи обмена веществ и энергии в организме
 Основным источником энергии для осуществления в организме процессов жизнедеятельности является биологическое окисление питательных веществ. На это окисление расходуется кислород. Следовательно, измерив количество потребленного организмом кислорода за минуту, час, сутки, можно судить о величине энергозатрат организма за время измерения.
Основным источником энергии для осуществления в организме процессов жизнедеятельности является биологическое окисление питательных веществ. На это окисление расходуется кислород. Следовательно, измерив количество потребленного организмом кислорода за минуту, час, сутки, можно судить о величине энергозатрат организма за время измерения.
Между количеством потребленного за единицу времени организмом кислорода и количеством образовавшегося в нем за это же время тепла существует связь, выражающаяся через калорический эквивалент кислорода (КЭО2). Под КЭО2 понимают количество тепла, образующегося в организме при потреблении им 1 л кислорода.
Способы оценки энергетических затрат организма. Прямая калориметрия основана на измерении количества тепла, непосредственно рассеянного организмом в теплоизолированной камере. При прямой калориметрии достигается высокая точность оценки энергозатрат организма, однако, ввиду громоздкости и сложности способ используется только для специальных целей.
462



 Непрямая калориметрия — основана на измерении количества потребленного организмом кислорода и последующем расчете энергозатрат с использованием данных о величинах дыхательного коэффициента (ДК) и КЭО2. Под дыхательным коэффициентом понимают отношение объема выделенного углекислого газа к объему поглощенного кислорода.
Непрямая калориметрия — основана на измерении количества потребленного организмом кислорода и последующем расчете энергозатрат с использованием данных о величинах дыхательного коэффициента (ДК) и КЭО2. Под дыхательным коэффициентом понимают отношение объема выделенного углекислого газа к объему поглощенного кислорода.
Сущность непрямой калориметрии видна на примере окисления глюкозы. Известно, что этот процесс описывается следующими превращениями: С6Н12О6 + 6О2 = 6СО2 + 6Н2О + 675ккал
При окислении 1 г глюкозы количество выделяющейся энергии составляет 675:180 (масса 1 моля глюкозы) = 3,75ккал. На окисление 1 моля глюкозы затрачивается 6 молей О2 или 134,4(6x22,4)л. Калорический эквивалент 1л О2, использованного на окисление глюкозы, равен 675ккал:134,4л = 5,02ккал/л. Так как смесь углеводов пищевых продуктов имеет несколько более высокую, чем чистая глюкоза, энергетическую ценность, то при окислении их в организме КЭО2 = 5,05ккал/л. Из приведенного уравнения реакции окисления глюкозы видно, что объем выделенного в процессе окисления углекислого газа равен объему затраченного кислорода. Следовательно, при окислении глюкозы
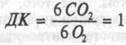
В случае окисления жиров, в которых на 1 атом углерода приходится меньше атомов кислорода, чем в углеводах и белках, величина ДК имеет значение 0,7. При окислении белковой, а также смешанной пищи величина ДК принимает промежуточное значение между 1,0 и 0,7. Поскольку в организме все питательные вещества одновременно подвергаются окислению, то определив величину ДК, можно условно судить о преимущественном окислении в организме того или иного вида питательных веществ. Так как для каждого питательного вещества характерна своя энергетическая ценность, то по величине дыхательного коэффициента можно рассчитать значение калорического эквивалента кислорода. Величины этих значений представлены в табл. 10.3.
Таблица 10.3 Значение ДК и КЭ при оксилении в организме различных питательных веществ
| Энергетическая ценность | О2 | СО2 | ДК | КЭО2 | |
| ккал/г | Л/ Г | Л/Г | ккал/л | ||
| Углеводы | 4,0 | 0,81 | 0,81 | 1,0 | 5,05 |
| Белки | 4,0 | 0,94 | 0,75 | 0,80 | 4,46 |
| Жиры | 9,0 | 1,96 | 1,39 | 0,70 | 4,69 |
463

 В условиях интенсивной физической нагрузки и при состояниях, когда в организме имеет место накопление углекислоты, ДК может принимать значение > 1,0. В этих случаях по нему нельзя судить о природе окисляемых веществ.
В условиях интенсивной физической нагрузки и при состояниях, когда в организме имеет место накопление углекислоты, ДК может принимать значение > 1,0. В этих случаях по нему нельзя судить о природе окисляемых веществ.








