 2015-03-22
2015-03-22 2256
2256Экскурсия 6. Ландшафтный профиль через боровой экологиче-ский ряд. Итак, наша задача — совершить экскурсию внутри одного и того же ландшафта, пытаясь объяснить и описать существующие внутрен-ние различия. Для ландшафтоведа в отличие от почвоведа или лесовода не столь уж важно скрупулезное описание почв и растительности по ди-агностическим признакам, сколько установление связей, причинно-след-ственных зависимостей между различными компонентами ландшафта. Поэтому в нашем описании мы будем останавливаться на главных при-знаках местообитания, которые могут быть выявлены в ходе полевых изысканий:
• положение в рельефе;
• механический состав верхней почвенной толщи;
• чередование разных по этому признаку слоев в профиле почв и грун та;
• уклон поверхности участка;
• интенсивность оподзоливания и наличие на той или иной глубине следов оглеения;
• мощность и структура лесной подстилки;
• состав и показатели продуктивности древесного яруса (класс бони тета, верхняя высота, запасы, годичный прирост стволовой массы);
• доминанты и субдоминанты нижних ярусов леса;
• наличие или отсутствие представителей тех или иных групп расте ний-индикаторов.
И все же главным диагностическим признаком для нас будет состав древостоев в коренных типах леса. По этому признаку биогеоценозы лес-ной зоны России могут быть «уложены» в три экологических ряда:
• боровой — в древостое господствует сосна;
• суборевый — наряду с сосной значительное участие принимает и ель;
• раменевый — древостой формируют ель.
Экологические ряды биогеоценозов — своего рода «периодическая таблица» ландшафтоведения (ее долгие годы разрабатывали и уточняли русские экологи П.С.Погребняк, В.Н.Сукачев, В.В.Алехин, А.Я.Орлов), позволившая увязать между собой взаимодействие и взаимозависимость различных ландшафтообразующих факторов.
Итак, попробуем разобраться — совершим экскурсию в ближайшие сосны на холмах. Нетрудно догадаться, что мы оказались в пределах вод-но-ледниковой (зандровой) равнины.
Начнем с верхушки холма. Здесь в погожий летний день сухо и сол-нечно. Невысокие тонкие стволы с негустыми кронами стоят неплотно, оставляя между собой окна и редины, куда свободно проникает свет, и где местами успешно развивается подрост, формируя быстро растущие группы жердняка.
В древостое заметны чуть более мощные экземпляры, но много и совсем сухих тонких стволов, которые способны повалиться от несиль-ного нажима.
Напочвенный покров в таком лесу весьма живописен: на фоне «окон» светло-желтого песка округлые пятна лишайников «исландского мха» (цетрария исландская) и «оленьего мха» (кладонии оленьей). Невероятно красивые подушечки лишайников напоминают игрушечные заросли, словно специально кем-то сделанные для домика лилипутов. (Кстати архитекторы зачастую используют лишайники именно в этом качестве, оформляя с помощью «кустиков» оленьего мха «ландшафт» на макетах городских микрорайонов). На самом деле лишайник — симбиоз двух ра-стений: водоросли и гриба, последний и прикрепляет «кустик» к песча-ному субстрату.
Травяной покров не развит: кроме лишайников в напочвенном покро-ве заметное участие могут принимать пятна зеленых мхов и небольшие куртины брусники. Единично встречается золотая розга, ландыш, плау-ны булавовидный и сплюснутый, овсяница овечья, кошачья лапка, марь-янник луговой.
Таким образом, перед нами самое сухое местообитание (или экотоп — буквально «место» в экологическом ряду) в пределах ландшафта. Выпа-дающие осадки долго не задерживаясь на вершине холма, быстро филь-труются через пески и скатываются по склонам. Верховодка всегда рас-положена глубоко (2 — 5 м от поверхности), следовательно, весь корне-обитаемый слой почвы (часть грунтов, в которых происходит почвообра-зование и располагается основная масса всасывающих корней сосны), находится выше зоны капиллярной каймы грунтовых вод — уровня, до которого могут подняться подземные воды в хороший теплый день за счет восходящего их движения по микротрубочками и трещинам почвы. Теперь попытаемся сделать небольшую в два-три штыка лопаты при-копку, поскольку процессами почвообразования в таком урочище затро-нута лишь самая верхняя часть почвогрунтов. Что же мы увидим? Разрез откроет нам однородные, тонкозернистые, как правило, хорошо сорти-рованные, т.е. перемытые или перевеянные, пески желтовато-коричне-вого, желтого или желтовато-бурого цвета, самый верхний горизонт ко-торых слегка осветлен. Это осветление можно в принципе считать при-знаками подзолистого процесса. Но только в принципе — у специалистов-
почвоведов нет уверенности по поводу сути процесса, вызывающего ос-ветление в данном экотопе. Полагают, что это может быть простая отмыв-ка пленок железистых и марганцевых оксидов с песчинок, поэтому цвет слоя определяется белесоватой окраской кварцевых частиц. Гумусовый горизонт на бедных песках тоже едва заметен, здесь нет того богатства органики, которое при разложении образует гумус.
Теперь, определив характер растительности и почв, мы можем назвать наш биогеоценоз: сосняк лишайниковый на скрытоподзолистых (или мелкоподзолистых) супесчаных почвах привершинных частей холмов водно-ледниковой равнины. Экотоп (местообитание), в котором сформи-ровался этот участок соснового леса, характеризуется двумя основными свойствами: бедностью толщи рыхлых кварцевых песков и сухостью, обусловленной положением в рельефе.
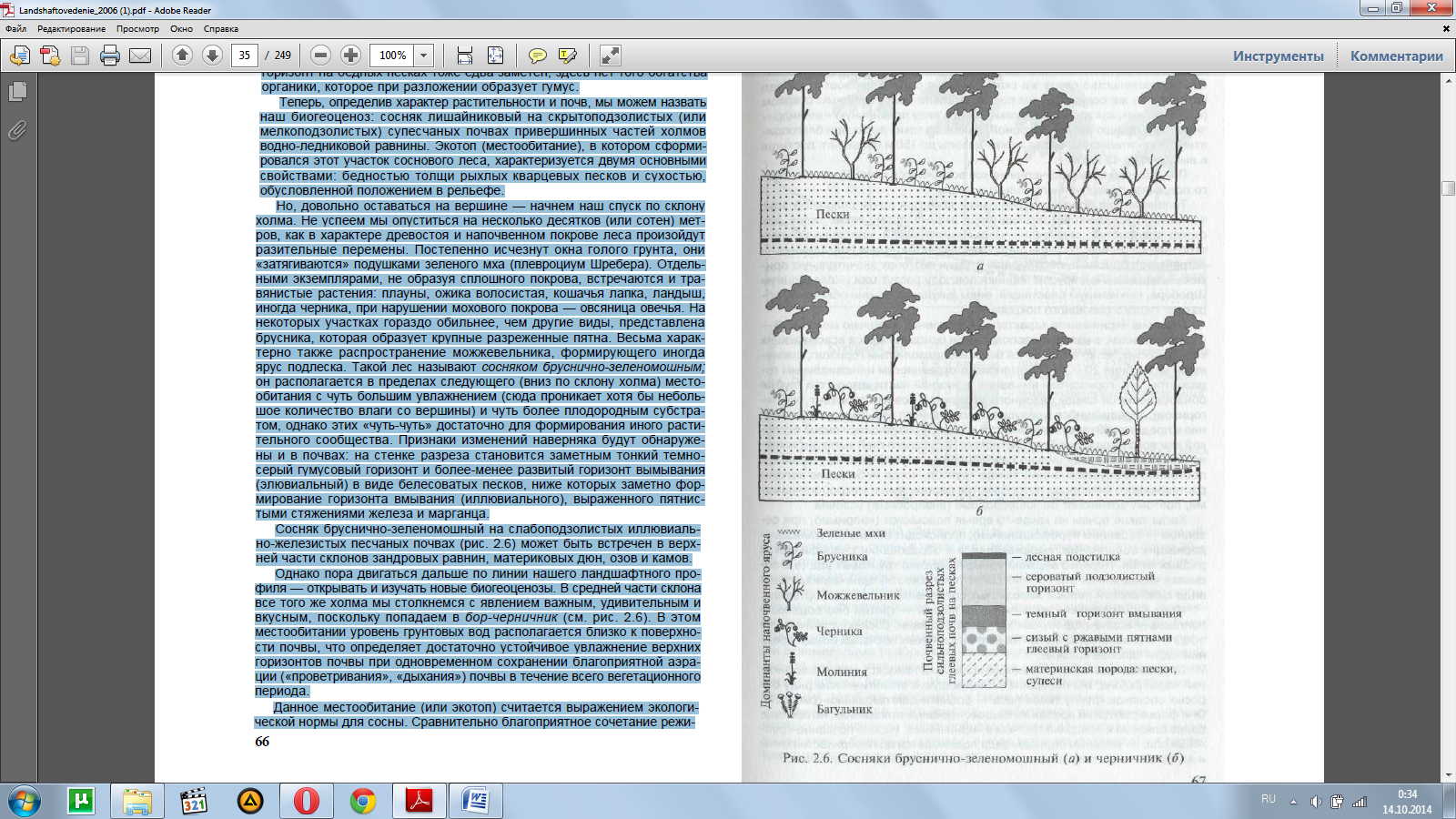
Но, довольно оставаться на вершине — начнем наш спуск по склону холма. Не успеем мы опуститься на несколько десятков (или сотен) мет-ров, как в характере древостоя и напочвенном покрове леса произойдут разительные перемены. Постепенно исчезнут окна голого грунта, они «затягиваются» подушками зеленого мха (плевроциум Шребера). Отдель-ными экземплярами, не образуя сплошного покрова, встречаются и тра-вянистые растения: плауны, ожика волосистая, кошачья лапка, ландыш, иногда черника, при нарушении мохового покрова — овсяница овечья. На некоторых участках гораздо обильнее, чем другие виды, представлена брусника, которая образует крупные разреженные пятна. Весьма харак-терно также распространение можжевельника, формирующего иногда ярус подлеска. Такой лес называют сосняком бруснично-зеленомошным; он располагается в пределах следующего (вниз по склону холма) место-обитания с чуть большим увлажнением (сюда проникает хотя бы неболь-шое количество влаги со вершины) и чуть более плодородным субстра-том, однако этих «чуть-чуть» достаточно для формирования иного расти-тельного сообщества. Признаки изменений наверняка будут обнаруже-ны и в почвах: на стенке разреза становится заметным тонкий темно-серый гумусовый горизонт и более-менее развитый горизонт вымывания (элювиальный) в виде белесоватых песков, ниже которых заметно фор-мирование горизонта вмывания (иллювиального), выраженного пятнис-тыми стяжениями железа и марганца.
Сосняк бруснично-зеленомошный на слабоподзолистых иллювиаль-но-железистых песчаных почвах (рис. 2.6) может быть встречен в верх-ней части склонов зандровых равнин, материковых дюн, озов и камов.
Однако пора двигаться дальше по линии нашего ландшафтного про-филя — открывать и изучать новые биогеоценозы. В средней части склона все того же холма мы столкнемся с явлением важным, удивительным и вкусным, поскольку попадаем в бор-черничник (см. рис. 2.6). В этом местообитании уровень грунтовых вод располагается близко к поверхно-сти почвы, что определяет достаточно устойчивое увлажнение верхних горизонтов почвы при одновременном сохранении благоприятной аэра-ции («проветривания», «дыхания») почвы в течение всего вегетационного периода.
Данное местообитание (или экотоп) считается выражением экологи-ческой нормы для сосны. Сравнительно благоприятное сочетание режи-
ма влажности и аэрации в черничниках обеспечивает достаточно успеш-ный рост малотребовательной к почвенным условиям скромницы-сосны, даже несмотря на потенциальную бедность рыхло-песчаной толщи почвы. Это обстоятельство сразу же сказывается в облике древостоя. Перед нами все та же сосна, но уже со значительно более мощным стволом («сбежистым», как говорят лесники, имея в виду правильную «цилиндри-чность») и хорошо развитой кроной. Именно в таких лесах при благопри-ятных обстоятельствах сосна может дожить до 150 и более лет, достигая в высоту 40 и 45 м.
Под пологом сосняков-черничников встречается очень мало сосново-го подроста, приуроченного только к окнам.
Типичным для боровых черничников является почти сплошной ярус черники с рассеянно встречающимися экземплярами таких видов, как ожика волосистая, седмичник, марьянник луговой, довольно обычны скопления плаунов. Чаще — на кочках и приствольных повышениях — встречается брусника, образующая в таких экотопах значительную при-месь к чернике. Под ярусом черники повсюду растут мхи — плевроциум Шребера, гилокомиум блестящий, виды дикранум, но они обычно не об-разуют густого сплошного покрова.
Для почв черничников характерно накопление довольно мощной (6 — 9 см) подстилки, в которой располагается основная масса всасывающих корней сосны; четко выделяется белесый подзолистый горизонт, сменя-емый на глубине 20 — 25 см интенсивно окрашенным иллювиальным го-ризонтом, т.е. горизонтом вмывания, в нижней части которого и глубже обнаруживаются следы сезонного оглеения. Глеевый горизонт почв — горизонт, образующийся в результате постоянного избыточного увлажне-ния в пределах 50 см от поверхности и характеризующийся сизой окрас-кой или ярко выраженной пятнистостью. Чаще всего глей выглядит имен-но как сизовато-серая сильно пачкающаяся «глина», формирующаяся в постоянно переувлажненной толще грунтов, когда небольшие запасы растворенного кислорода потребляются растениями и микроорганизма-ми, поэтому возникают бескислородные (анаэробные) условия.
Когда такие почвы на какое-то время подсыхают (например, при се-зонном — весеннем переувлажнении), происходит окисление железосо-держащих соединений, выражающееся в образовании специфических ржавых пятен. Нередко верхний уровень зеркала грунтовых вод (а в со-сняках черничниках это 30 — 40 см от поверхности) запечатлевается в виде красноватой полосы железистых соединений — корочки ортзанда.
Итак, на «ступеньке» экологической нормы — третий биогеоценоз в ландшафте водно-ледниковых холмистых равнин: сосняк-черничник на средне- и сильноподзолистых глееватых и глеевых песчаных почвах сред-ней части склонов холмов.
Спускаясь еще ниже к подножью холма или находясь в пределах ниж-ней части склона, мы попадаем в следующую в экологическом ряду бо-ровых сосняков группу типов леса — сосняки-долгомошники (рис. 2.7). Они формируются на все тех же бедных песчаных отложениях, но при еще более близком к поверхности, чем в черничниках, уровне почвенно-грун-товых вод; во влажные периоды вода поднимается до поверхности почвы и анаэробные условия периодически охватывают всю толщу почвы.

Рис. 2.7. Сосняк-долгомошник
Ухудшение условий обитания сразу же сказывается на внешнем об-лике деревьев — сосны в таком лесу редко достигают высоты более 20 — 25 м, ствол их уже не отличается «сбежистостью»: на нем заметны утол-щения вокруг годовых мутовок, наросты и искривления.
«Долгий мох», или кукушкин лен (рис. 2.8), хорошо узнается по «коро-бочкам» на концах, образует сплошной, очень мощный покров, сменяю-щийся лишь в микропонижениях сфагновыми мхами, а на приствольных повышениях зелеными мхами. Среди покрова кукушкина льна единичны-ми экземплярами разбросаны ожика волосистая, марьянник луговой, лесной вейник, ятрышник, щучка, осока шаровидная.
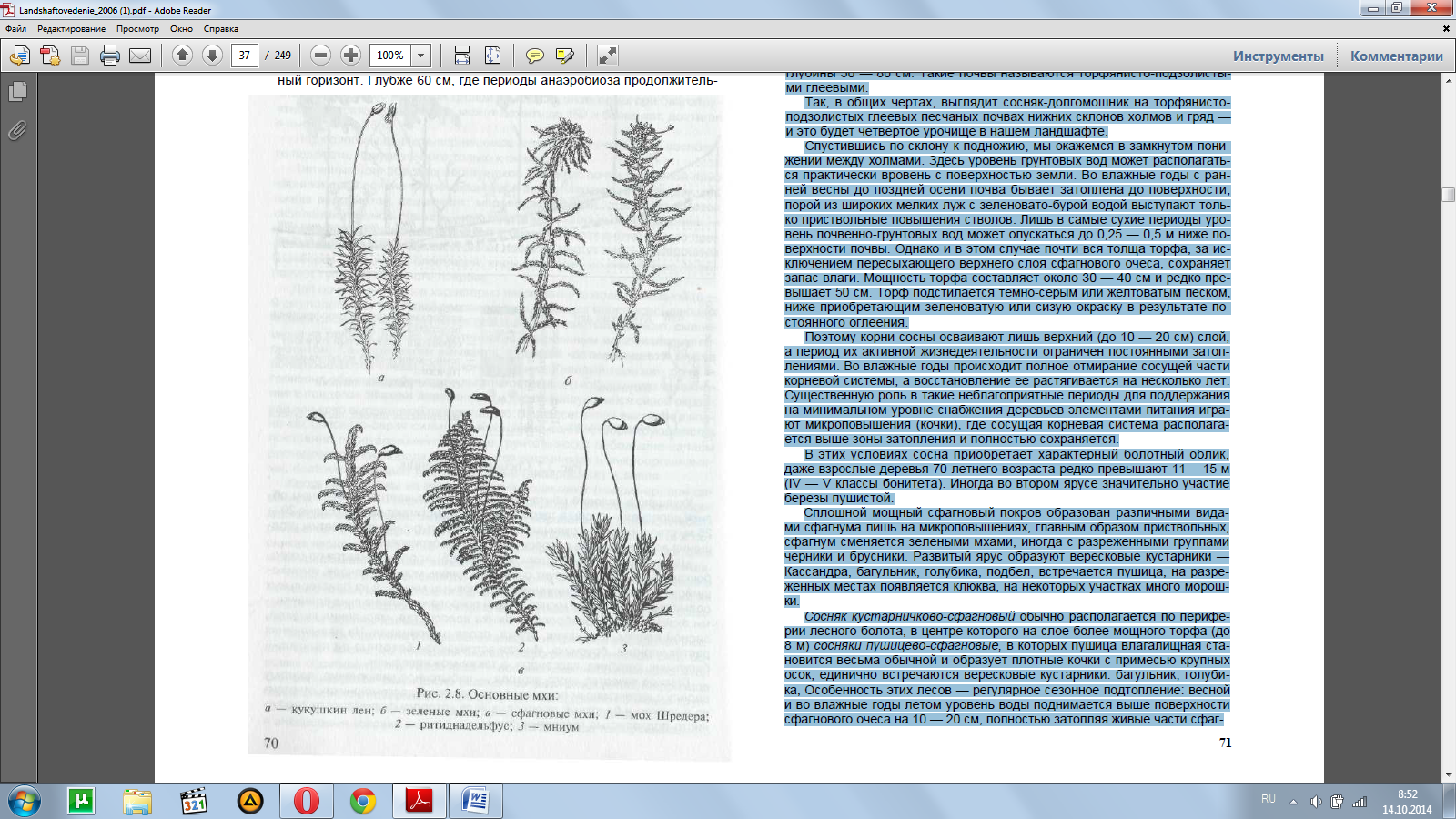
На повышениях растет черника, брусника. Иногда встречаются болотные кустарники — багульник, голубика, Кассандра, а также можжевельник.
Главное качество этого экотопа — избыточное увлажнение, поэтому наряду с интенсивным подзолообразованием в долгомошниках отчетли-во выражен и болотный процесс, в ходе которого формируются торфя-нисто-подзолистые глееватые почвы.
Как выглядят такие почвы? Грубая подстилка из остатков кукушкина льна и хвои достигает толщины 10—12 см. Продукты неполного разло-жения органических веществ окрашивают подзолистый горизонт в се-роватый цвет и проникают глубже, формируя очень типичный для дол-гомошников коричневый или темно-коричневый гумусово-иллювиаль-ный горизонт. Глубже 60 см, где периоды анаэробиоза продолжитель-ны, появляется пятнистость: чередование зеленовато-сизых и ярко-ржавых и бурых пятен или слоев, а на глубине около 80 см обычно рас-полагается сплошной сизый горизонт глея. Сосущие корни приурочены в основном к подстилке, хотя единичные корни встречаются иногда и до глубины 50 — 80 см. Такие почвы называются торфянисто-подзолисты-ми глеевыми.
Так, в общих чертах, выглядит сосняк-долгомошник на торфянисто-подзолистых глеевых песчаных почвах нижних склонов холмов и гряд — и это будет четвертое урочище в нашем ландшафте.
Спустившись по склону к подножию, мы окажемся в замкнутом пони-жении между холмами. Здесь уровень грунтовых вод может располагать-ся практически вровень с поверхностью земли. Во влажные годы с ран-ней весны до поздней осени почва бывает затоплена до поверхности, порой из широких мелких луж с зеленовато-бурой водой выступают толь-ко приствольные повышения стволов. Лишь в самые сухие периоды уро-вень почвенно-грунтовых вод может опускаться до 0,25 — 0,5 м ниже по-верхности почвы. Однако и в этом случае почти вся толща торфа, за ис-ключением пересыхающего верхнего слоя сфагнового очеса, сохраняет запас влаги. Мощность торфа составляет около 30 — 40 см и редко пре-вышает 50 см. Торф подстилается темно-серым или желтоватым песком, ниже приобретающим зеленоватую или сизую окраску в результате по-стоянного оглеения.
Поэтому корни сосны осваивают лишь верхний (до 10 — 20 см) слой, а период их активной жизнедеятельности ограничен постоянными затоп-лениями. Во влажные годы происходит полное отмирание сосущей части корневой системы, а восстановление ее растягивается на несколько лет. Существенную роль в такие неблагоприятные периоды для поддержания на минимальном уровне снабжения деревьев элементами питания игра-ют микроповышения (кочки), где сосущая корневая система располага-ется выше зоны затопления и полностью сохраняется.
В этих условиях сосна приобретает характерный болотный облик, даже взрослые деревья 70-летнего возраста редко превышают 11 —15 м (IV — V классы бонитета). Иногда во втором ярусе значительно участие березы пушистой.
Сплошной мощный сфагновый покров образован различными вида-ми сфагнума лишь на микроповышениях, главным образом приствольных, сфагнум сменяется зелеными мхами, иногда с разреженными группами черники и брусники. Развитый ярус образуют вересковые кустарники — Кассандра, багульник, голубика, подбел, встречается пушица, на разре-женных местах появляется клюква, на некоторых участках много морош-ки.
Сосняк кустарничково-сфагновый обычно располагается по перифе-рии лесного болота, в центре которого на слое более мощного торфа (до 8 м) сосняки пушицево-сфагновые, в которых пушица влагалищная ста-новится весьма обычной и образует плотные кочки с примесью крупных осок; единично встречаются вересковые кустарники: багульник, голуби-ка, Особенность этих лесов — регулярное сезонное подтопление: весной и во влажные годы летом уровень воды поднимается выше поверхности сфагнового очеса на 10 — 20 см, полностью затопляя живые части сфаг-новых мхов. Почва может оставаться затопленной в течение почти всего периода вегетации. В периоды сухих и средних по увлажнению лет корни сосны обильно насыщают верхний (до 10 см) слой торфа, проникая до глубины 25 см. Но в очень влажные годы происходит полное отмирание всех сосущих и мелких проводящих корней сосны и березы. Сохраняют-ся лишь скелетные корни (толще 1 —2 см) и все корни на высоких мик-роповышениях, к которым и приурочены деревья. Сосна пушицево-сфаг-новых сосняков выглядит совсем карликовой, не более 4 — 8 м в высоту.
Итак, пятый биогеоценоз в нашем экологическом ряду — сосняк сфаг-новый (кустарничково-сфагновый или пушицево-сфагновый) на торфяных почвах в межхолмовых понижениях и замкнутых котловинах.
Таким образом, находясь в пределах одного и того же ландшафта, в одной лишь боровой экологической линии, мы обнаруживаем пять (!) экотопов с соответствующими биогеоценозами (рис. 2.9).

2.4. Суборевый экологический ряд — елово-сосновые леса
Оставим песчаные пространства водно-ледниковой равнины и выберемся на поверхность более сложного литологического стро-ения — озерно-ледниковую равнину, где в составе отложений наряду с песками участвуют заметные прослойки суглинков. В та-ких условиях развиваются биогеоценозы «подборового», или, как его принято называть, суборевого факторально-экологического ряда. В суборевом ряду (рис. 2.10) наряду с фактором увлажнения на дифференциацию ландшафтов сильное влияние оказывает фак-тор трофности («питательности») субстрата, обусловленный на-личием суглинистых слоев в песчаной толще.

Присутствие даже небольшой глинистой прослойки в пределах корнеобитаемого слоя уже дает примесь ели к сосне, поэтому лесные сообщества этого типа и получили в лесоведении название «суборь».
Интересно, что фактор трофности грунтов становится веду-щим в дифференциации лесных ландшафтов при нейтрализации фактора увлажнения, т.е. в условиях «нормы»: на средних частях пологих склонов озерно-ледниковых равнин, плоских поверхно-стях надпойменных террас и т.д. И, наоборот, в крайних (по шка-ле увлажнения) условиях различия между лесными сообществами борового и суборевого ряда становятся не слишком заметными: растения оказываются не в состоянии взять из почвы азот и дру-гие элементы питания при неблагоприятном водно-воздушном режиме. В суборевом ряду нет группы лишайниковых сосняков, так как присутствие более влагоемких суглинистых прослоек не создает абсолютно сухого водного режима, обусловливающего формиро-вание сообществ этой группы. В «сухой» части суборевого ряда — мшистые сосняки с елью и сосняки-брусничники с елью на подзоли-стых супесчано-суглинистых почвах (рис. 2.11).

Гумусовый гори-зонт почв в этих лесах мощнее и лучше выражен (более темный) чем аналогичном чистых сосняках.
В мшистых суборевых сосняках формируются высокополнот-ные древостой I и II классов бонитета с чисто сосновым (или при участии березы поникшей) первым ярусом. Во втором ярусе зна-чительно усиливаются позиции ели. В мало измененных рубками или пожарами лесах ель образует ярус подроста и тонкомера с отдельными деревьями, выходящими в первый ярус, но не до-стигающими размеров сосны. Сплошной моховой покров образован, главным образом, плев-роциумом Шребера, в окнах иногда встречаются изреженные пят-на лишайников. Травяно-кустарничковый покров обычно не вы-ражен: единично отмечаются плауны, ожика волосистая, орти-лия однобокая, грушанка круглолистная, кошачья лапка, золо-тая розга, ландыш, черника. В отличие от боровой группы на некоторых участках можно встретить индикатор более плодород-ных (по сравнению с боровыми) почв — кислицу, а также лин-нею северную, седмичник. Практически всегда единичными груп-пами присутствует брусника, реже образуя выраженный сплош-ной ярус.
Сравнительно маломощная подстилка не препятствует возоб-новлению хвойных под пологом леса, и там, где нет ели, под пологом встречается иногда обильный подрост сосны. Но он угне-тен, периодически отмирает, заменяясь новыми генерациями, и лишь в окнах формирует более или менее быстрорастущие группы. Ель серьезно затрудняет возобновление сосны, но не может пол-ностью его подавить, даже на тех участках, где длительное время не было пожаров. Ель не образует здесь сплошного сомкнутого полога, кроны ее не густые, она менее долговечна, во время силь-ных засух угнетается еще сильнее и выпадает.
Очень близки к соснякам мшистым и брусничникам сосняки орляковые (с господством папоротника орляка, образующего плот-ные заросли), отличаясь от них чуть большей питательностью суб-страта.
Ниже по рельефу в условиях нормального увлажнения в субо-ревом ряду — сосняки черничные с елью. Они получают развитие там, где почвообразующие породы представлены супесями, пы-леватыми песками либо в профиле почвы встречаются суглини-стые прослойки или линзы. На глубине около 0,5—1,0 м часто подстилаются суглинками.
Сосновые древостой в таких условиях развиваются по I классу бонитета — это очень красивые леса и дополнительную красоту им придает участие ели, кроны которой располагаются главным образом во втором ярусе, но отдельные «готические» остроконе-чные вершины входят и в господствующий сосновый полог дре-востоев. Улучшение режима питания в суборевых черничниках по сравнению с боровыми усиливает в этих достаточно благоприят-ных условиях увлажнения позиции березы поникшей. Состав фор-мирующихся на открытых местах древостоев, очевидно, зависит от того, какая из этих двух светолюбивых и примерно одинаково быстро растущих пород раньше займет свободную территорию. Если осеменение сосной произойдет на несколько лет позднее, чем березы, то береза, опередив в росте сосну, будет ее угнетать и сосна в большей или меньшей мере полностью выпадет из соста-ва древостоя. В других случаях сосна опережает березу, которая, попав под сосновый полог, растет медленно, постепенно погиба-ет или сохраняется только во втором ярусе.
По составу и развитию доминант нижних ярусов сообществ боровые и суборевые черничники довольно близки. Основной до-минантой в обоих случаях является черника. Но улучшение плодо-родия почв способствует появлению или более широкому уча-стию в суборевых экотопах требовательных к условиям питания видов: в подлеске — крушины ломкой, рябины; в наземном по-крове — майника, костяники, орляка, седмичника, вейника лес-ного, который так же, как и орляк, встречается иногда довольно обильно, образуя разреженный ярус над черникой. Такой биогео-ценоз называется лес елово-сосновый черничный, или чернично-сфаг-новый (с примесью березы) на подзолистых супесчаных или супесча-но-суглинистых почвах (рис. 2.12).

Ниже по рельефу располагается экотоп долгомошной суборевой группы; леса этой группы сходны с описанными выше чистыми долгомошниками. Однако существенным отличием экотопа слу-жит наличие на небольшой глубине (40 — 60 см) суглинистых или супесчано-суглинистых прослоек и включений.
Кустарничково-сфагновая и пушицево-сфагновая суборевые группы формируются на торфянисто-подзолистых, подзолисто-гле-евых супесчаных почвах на поверхности плоских бессточных котло-вин, в нижней части склонов. Они отличаются также составом по-чвообразующих пород: пески или супеси, подстилаемые суглинка-ми, иногда с поверхности перекрытые маломощным слоем торфа.
Кустарничково-сфагновые сообщества формируются и на отло-жениях озерного происхождения, хорошо изученных по данным пыльцевого анализа и стратиграфии торфяной залежи.
Леса в заболоченных низинах, подпитываемых грунтовыми во-дами, проходят одни и те же стадии эволюции, связанные с умень-шением доступных элементов питания (азота, фосфора, калия и др.). В начальной стадии существование низинного болота связано с травянистой влаголюбивой растительностью, которая со време-нем сменяется эвтрофными заболоченными лесами (сосняками, ольшаниками). Эти леса по мере нарастания толщи торфа уступа-ют место мезотрофным заболоченным лесам (соснякам, березня-кам), которые, в свою очередь, сменяются сфагновыми олигот-рофными сосновыми лесами.
Подобные экосистемы формируются и на песчаных отложени-ях высоких речных террас в их тыловой пониженной части, и в пределах зандровых равнин, там, где гидрологический режим обус-ловливает постоянно высокий уровень грунтовых вод, продолжи-тельное подтопление и нарастание толщи торфа.
2.5. Лесные биогеоценозы на суглинистых почвах
(рамени)
Рамени — старинное слово, обозначающее еловые, темнохвой-ные леса. Коренные ельники Русской равнины образуют несколь-ко экологических групп, занимающих разные экотопы в пределах моренных равнин и конечно-моренных возвышенностей с преоб-ладанием моренных валунных суглинков по всей верхней толще. Соответственно верхние корнеобитаемые горизонты почв в ко-ренных ельниках по механическому составу относятся обычно к легким суглинкам, реже к средним суглинкам или глинам.
Представление о «сосновых» и «еловых» почвах бытовало в на-роде еще с давних времен, но лишь в XX в. в связи с развитием геохимии ландшафта были выяснены действительные причины почвенных предпочтений сосны и ели. Выяснилось, что ель фор-
мирует чистые древостой, вытесняя сосну лишь при достаточно благоприятном режиме кальция в почве, причем не только на дренированных, но и на переувлажненных почвах с подтоком вод, обогащенных карбонатами кальция и магния.
По этой же причине ель совсем неплохо себя чувствует и на тяжело супесчаных почвах конечных морен, обогащенных глини-стыми частицами и рухляком обломочных пород. Однако и сосна при прочих благоприятных условиях может заходить на суглини-стые почвы. Мы отмечаем это обстоятельство отнюдь не затем, чтобы окончательно запутать начинающих ландшафтоведов, а лишь для того, чтобы подчеркнуть: в природе нет твердых и жестких зависимостей между любыми двумя парами факторов (например, в паре механический состав почв — доминанта лесного биогеоце-ноза). Именно этим и объясняется отсутствие резких переходов и границ между растительными сообществами — во-первых, и чрез-вычайное разнообразие лесных ландшафтов — во-вторых.
Кроме того, распространению еловых лесов во многом сопут-ствует неглубокое залегание карбонатной морены, которая очень широко распространена в центральной и восточной частях юж-ной тайги.
Хотя в целом для раменевого экологического ряда в пределах лесной зоны характерны суглинистые почвы, строение почвен-ного профиля может быть разнообразным. В центральной части южной тайги чаще встречаются двучленные почвы с верхней лег-косуглинистой или супесчаной толщей, подстилаемой на глубине 50 — 80 см обычно моренным валунным суглинком. Глубина зале-гания тяжелого суглинка определяет «шансы» сосны участвовать в составе еловых древостоев.
В зону произрастания ельников попадают и озерно-леднико-вые равнины, и надпойменные террасы, и широкие озерные низ-менности, поскольку в пределах всех этих поверхностей мы также сталкиваемся с залеганием более легких по механическому составу верхних слоев на расположенных глубже суглинках — озерных, аллювиальных и т.д.
В силу экологических особенностей ели раменевая группа лише-на «сухого» местообитания, который был бы аналогичен лишай-никовым борам, и начинается сразу с центрального члена ряда, представляющего экологическую «норму» — ельника-кисличника. Кислица очень теневынослива, и поэтому она одна способна фор-мировать негустой покров по затененным местам в сомкнутых груп-пах деревьев и подроста или под кронами крупных елей.
Ельники-кисличники (рис. 2.13) развиваются на своеобразных слабоподзолистых суглинистых почвах, с нечетким разделением элювиального горизонта (А) и горизонта вмывания (В).
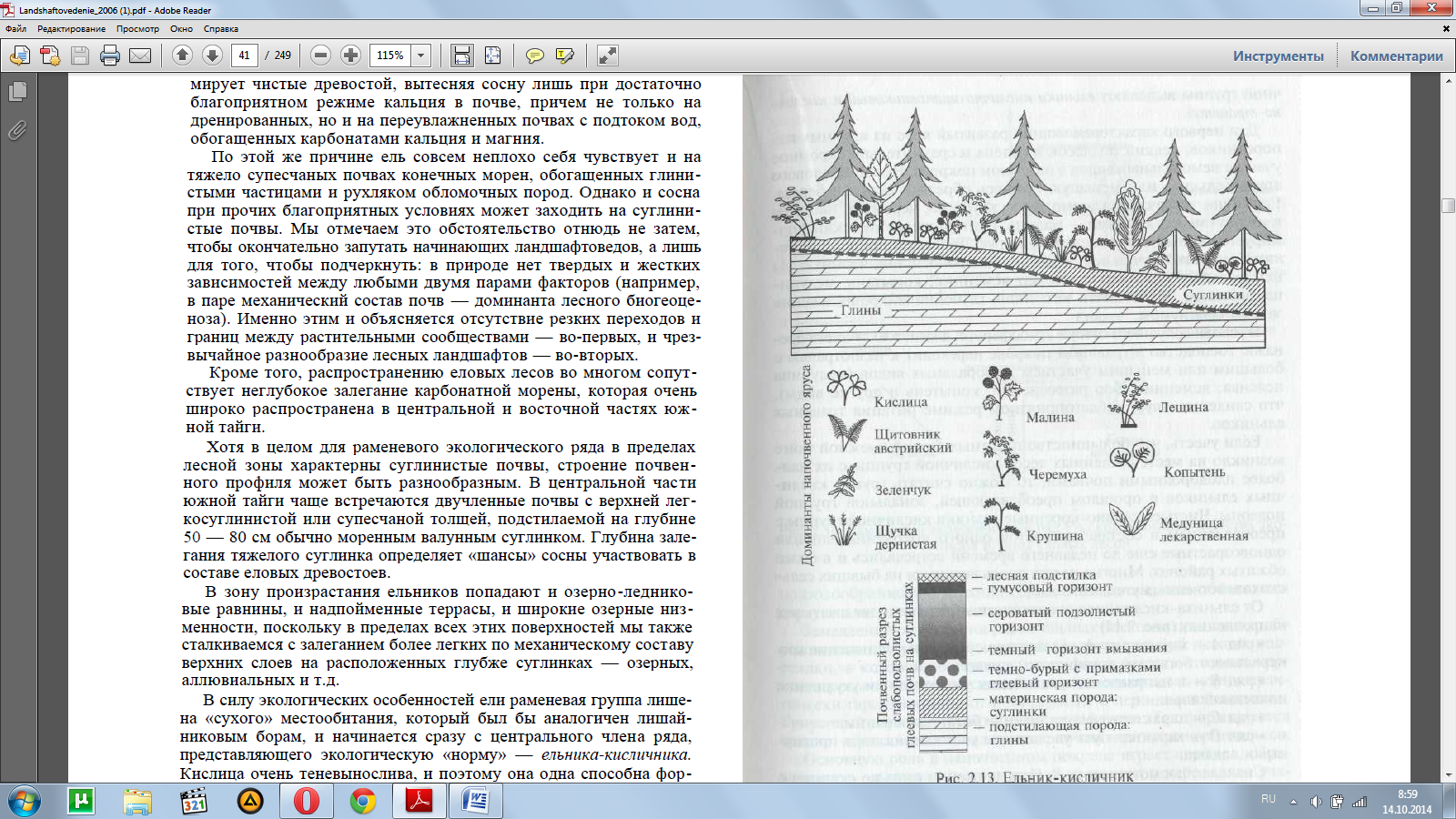
На глуби-не около 60 — 70 см залегает водоупорный тяжелый моренный суг-линок, над которым во влажные годы длительно задерживаетсяверховодка и развиваются анаэробные процессы, вызывающие оглеение. Следы бескислородного режима хорошо заметны: не-большие белесые пятна, марганцево-железистые мелкие конкре-ции. В связи с этим скелетная корневая система ели поверхност-ная и деревья в таких урочищах подвержены ветровалу.
В районах конечных морен и других возвышенных территорий преобладают леса кисличной группы, поэтому в пределах кисли-чной группы выделяют ельники кислично-щитовниковый и кислич-но-травяной.
Для первого характерен мощно развитый ярус из крупных па-поротников, редкий подлесок из клена и сравнительно скромное участие неморальных видов в наземном покрове. В составе елового яруса большую или меньшую примесь образуют осина и береза. Единично встречаются жимолость, крушина, черемуха, лещина; в «окнах» между кронами — малина. Щитовник австрийский фор-мирует мощный почти сплошной ярус. Травяной покров — кис-лица, майник, щучка дернистая, ожика волосистая, черника. Из неморальных видов присутствуют ясменник, копытень, медуница, зеленчук. Мхов почти нет, лишь глубокие микропонижения заняты сфагнумом.
В условиях лучшего оттока атмосферной влаги, т. е. лучшего дре-нажа, господство в травяном покрове переходит к разнотравью с большим или меньшим участием неморальных видов (медуница неясная, ясменник, бор развесистый, копытень и другие виды), что свидетельствует о благоприятном режиме питания травяных ельников.
Если учесть, что большинство пахотных угодий в южной тайге возникло на месте сведенных лесов кисличной группы с их наи-более плодородными почвами, то можно считать группу кисли-чных ельников в прошлом преобладающей, зональной группой подзоны. Чистые условно-коренные ельники кисличной группы с преобладанием в составе древостоя одного поколения ели или одновозрастные еще до недавнего времени встречались и в более обжитых районах. Многие из них вновь возникли на бывших сель-скохозяйственных угодьях.
От ельника-кисличника экологические ряды отходят в четырех направлениях (рис. 2.14):
• ряд А — характеризует увеличение сухости и уменьшение ми нерального богатства (трофности) почвы;
• ряд В — в направлении увеличения увлажнения и ухудшение почвенной аэрации;
• ряд С — характеризует увеличение богатства почв;
• ряд D — характеризует увеличение увлажнения почв проточ ными водами.

Сначала просмотрим ряд В. При движении вниз по склонам с увеличением увлажнения и одновременным ухудшения условий «дыхания» почв формируются урочища ельников-черничников (рис. 2.15).
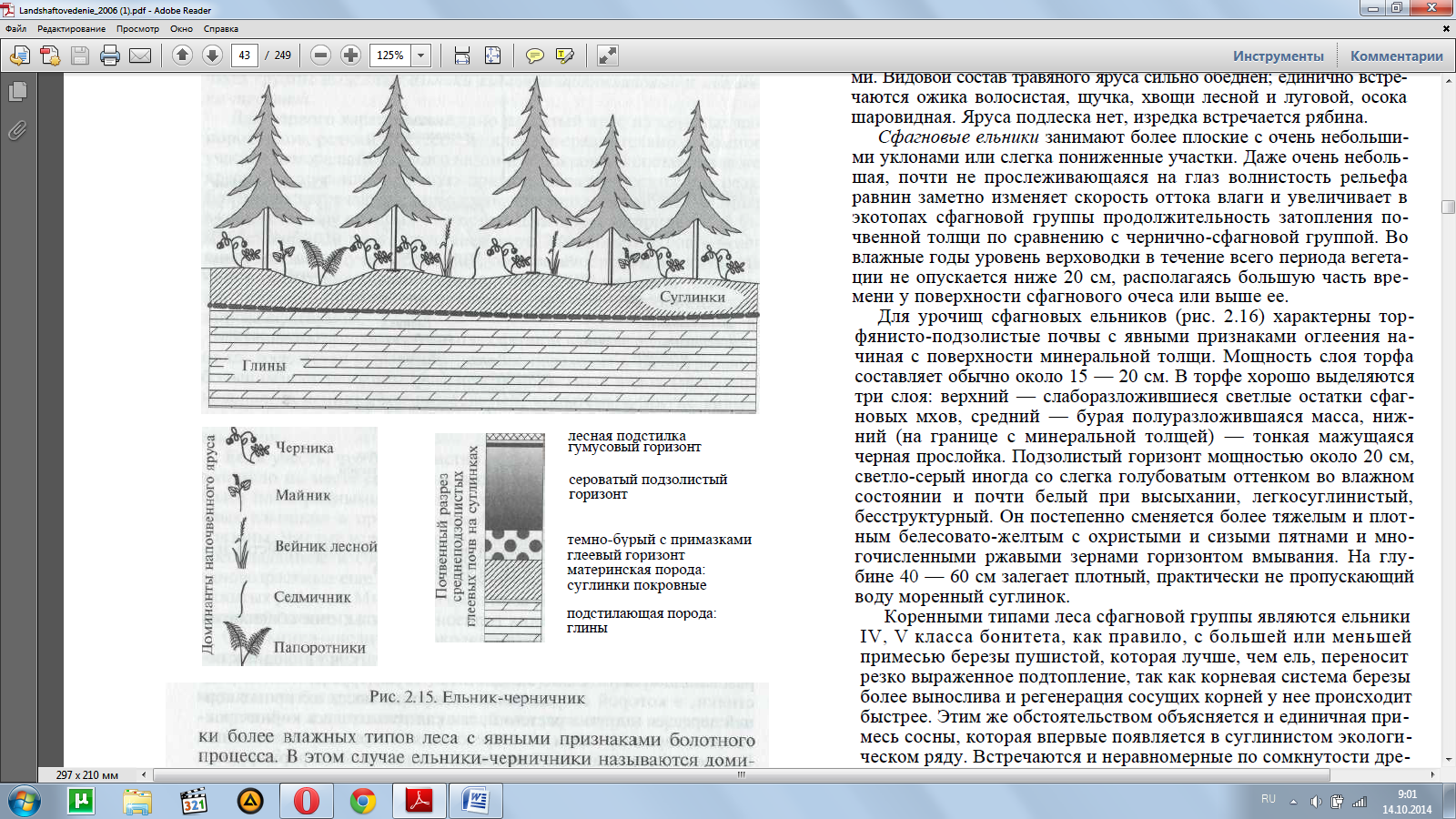
На более пологих склонах моренных равнин замедлен-ный отток влаги обусловливает подъем верховодки и периодичес-кое (весной, а во влажные годы и летом) затопление почвы до ее поверхности. Эта важнейшая особенность предопределяет направ-ленность почвообразования: при разложении растительных остат-ков возникают более кислые подвижные продукты, усиливаетсяподзолообразование верхних горизонтов почвы, они обедняются доступными элементами питания.
Замедление разложения растительных остатков приводит к об-разованию сравнительно мощной (3 — 6 см) грубой лесной под-стилки, в которой сосредоточена основная масса активных кор-ней деревьев и других растений, так как только здесь корни прак-тически гарантированы от повреждения в периоды подтопления. Гумусовый горизонт практически не выражен. Так формируются средне- и реже сильноподзолистые почвы ельников-черничников.
Основную роль в напочвенном покрове играет черника, обра-зующая в ельниках обычно сплошной покров с небольшим уча-стием вейника лесного, майника, седмичника, мелких папорот-ников и других видов таежного мелкотравья.
Волнистость и беспорядочная бугристость рельефа моренных равнин определяет мозаичность в распределении лесных урочищ, чутко реагирующих на изменение комплекса почвенных факто-ров, прежде всего режима аэрации и питания. Поэтому среди пре-обладающих по площади ельников-кисличников и ельников-чер-ничников вкраплены по плоским поверхностям и низинам участ-
ми. Видовой состав травяного яруса сильно обеднен; единично встре-чаются ожика волосистая, щучка, хвощи лесной и луговой, осока шаровидная. Яруса подлеска нет, изредка встречается рябина.
Сфагновые ельники занимают более плоские с очень небольши-ми уклонами или слегка пониженные участки. Даже очень неболь-шая, почти не прослеживающаяся на глаз волнистость рельефа равнин заметно изменяет скорость оттока влаги и увеличивает в экотопах сфагновой группы продолжительность затопления по-чвенной толщи по сравнению с чернично-сфагновой группой. Во влажные годы уровень верховодки в течение всего периода вегета-ции не опускается ниже 20 см, располагаясь большую часть вре-мени у поверхности сфагнового очеса или выше ее.
Для урочищ сфагновых ельников (рис. 2.16) характерны тор-фянисто-подзолистые почвы с явными признаками оглеения на-чиная с поверхности минеральной толщи. Мощность слоя торфа составляет обычно около 15 — 20 см. В торфе хорошо выделяются три слоя: верхний — слаборазложившиеся светлые остатки сфаг-новых мхов, средний — бурая полуразложившаяся масса, ниж-ний (на границе с минеральной толщей) — тонкая мажущаяся черная прослойка. Подзолистый горизонт мощностью около 20 см, светло-серый иногда со слегка голубоватым оттенком во влажном состоянии и почти белый при высыхании, легкосуглинистый, бесструктурный. Он постепенно сменяется более тяжелым и плот-ным белесовато-желтым с охристыми и сизыми пятнами и мно-гочисленными ржавыми зернами горизонтом вмывания. На глу-бине 40 — 60 см залегает плотный, практически не пропускающий воду моренный суглинок.
Коренными типами леса сфагновой группы являются ельники IV, V класса бонитета, как правило, с большей или меньшей примесью березы пушистой, которая лучше, чем ель, переносит резко выраженное подтопление, так как корневая система березы более вынослива и регенерация сосущих корней у нее происходит быстрее. Этим же обстоятельством объясняется и единичная при-месь сосны, которая впервые появляется в суглинистом экологи-ческом ряду. Встречаются и неравномерные по сомкнутости дре-востой с большими лишенными деревьев прогалинами и совсем чахлыми елками. В напочвенном покрове господствует сфагновые мхи, часто с большим участием кукушкина льна; совместно они образуют живописный желто-зеленый мозаичный покров. На при-стволовых возвышениях, гниющем валеже, крупных кочках рас-тут зеленые мхи. Сюда же приурочено и большинство растений очень бедного по видовому составу травяного покрова: марьян-ник луговой, щучка, брусника, золотая розга, седмичник, вей-ники лесной и Лангсдорфа, иван-чай. На ровных и пониженных местах по покрову мхов встречаются лишь единичные растения ситника развесистого, хвоща лесного.

Рис. 2.16. Ельник сфагновый
В очень изреженном подлеске встречаются рябина; ивы уша-стая и пепельная по наиболее переувлажненным участкам (где березовый древостой изрежен) образуют довольно густой подле-сок высотой 1,0— 1,5 м. Елового подроста мало, и распределен он очень неравномерно, располагаясь, главным образом, на микро-повышениях.
Ельники сфагновые почти не сохранились в южной тайге, встре-чаются лишь в более глухих районах.
Урочища сфагновой группы на суглинистых торфянисто-гле-евых почвах можно рассматривать как начальный этап болотооб-разования, который в дальнейшем, по мере нарастания торфа, неизбежно приведет к еще большему переувлажнению, уменьше-нию трофности почвогрунтов и к смене ельников заболоченными сосняками, а затем и сфагновыми болотами.
Итак, мы рассмотрели типы раменевых урочищ суглинистых отложений равнинной тайги, для которых характерно увлажне-ние за счет выпадающих непосредственно на данную площадь ат-мосферных осадков, а отток влаги происходит, главным образом, по начальным элементам гидрографической сети — ложбинам и лощинам, временным водостокам, заполняющимся лишь талой водой или после обильных дождей.
Но существует весьма разнообразные экотопы, которым свой-ственно присутствие влаги, перемещающейся с соседних участ-ков внутри или по поверхности почвы и обогащенной питатель-ными элементами. Рельеф тяжелосуглинистых отложений основ-ной морены осложнен малозаметными узкими или довольно ши-рокими корытоообразными ложбинами, по которым идет сток вод весной и при обильных летних дождях. Эти ложбины в низо-вых частях сливаются друг с другом, и при пологом рельефе могут формироваться более или менее широкие полосы, а в низинах и значительные участки биогеоценозов проточного увлажнения с зарослями таволги, с болотным высокотравьем или покровом из сфагновых мхов.
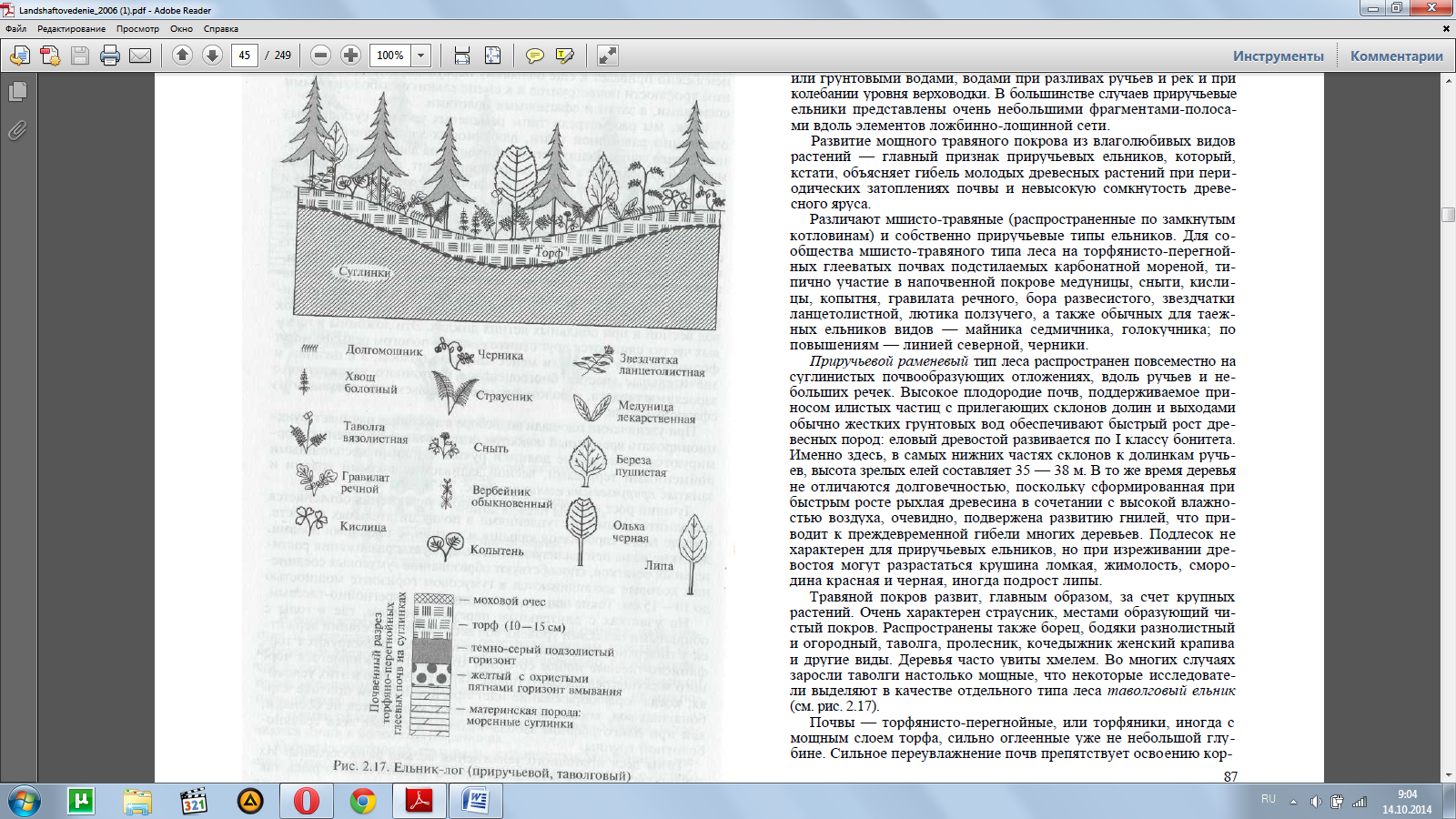
При увеличении площади водосбора в ложбинах начинает функ-ционировать временный водосток, а затем ниже по течению фор-мируются выраженные долинки ручьев с узкими несплошными пойменными террасами, весной заливаемые полыми водами и занятые приручьевыми ельниками (рис. 2.17).
Лучший рост в типах леса проточного увлажнения объясняется дополнительными поступлениями в почву питательных веществ, прежде всего карбонатов кальция и магния, с текущими водами. Жесткие воды нейтрализуют кислые продукты разложения расти-тельных остатков, способствуют образованию гумусовых соедине-ний которые накапливаются в гумусовом горизонте мощностью до 10—15 см. Такие почвы можно отнести к перегнойно-глеевым. На участках с длительным переувлажнением, где в годы с обильными осадками уровень вод весь период вегетации держится у поверхности почвы, а часто и выше ее, формируются тор-фянисто-глеевые почвы со слоем хорошо разложившегося чер-ного мажущегося рыхлого торфа. Характерно, что в этих услови-ях, когда торфообразование идет при постоянном притоке кар-бонатных вод, коренной растительностью являются не сосняки, как при олиготрофном заболачивании, а еловые леса травяно-болотной группы.
Типы леса проточного увлажнения весьма разнообразны. Их трудно расположить в пределах одного экологического ряда, так
как основная особенность — увлажнение обогащенными минера-лами водами — достигается разными путями: с поверхностными или грунтовыми водами, водами при разливах ручьев и рек и при колебании уровня верховодки. В большинстве случаев приручьевые ельники представлены очень небольшими фрагментами-полоса-ми вдоль элементов ложбинно-лощинной сети.
Развитие мощного травяного покрова из влаголюбивых видов растений — главный признак приручьевых ельников, который, кстати, объясняет гибель молодых древесных растений при пери-одических затоплениях почвы и невысокую сомкнутость древе-сного яруса.
Различают мшисто-травяные (распространенные по замкнутым котловинам) и собственно приручьевые типы ельников. Для со-общества мшисто-травяного типа леса на торфянисто-перегной-ных глееватых почвах подстилаемых карбонатной мореной, ти-пично участие в напочвенной покрове медуницы, сныти, кисли-цы, копытня, гравилата речного, бора развесистого, звездчатки ланцетолистной, лютика ползучего, а также обычных для таеж-ных ельников видов — майника седмичника, голокучника; по повышениям — линией северной, черники.
Приручьевой раменевый тип леса распространен повсеместно на суглинистых почвообразующих отложениях, вдоль ручьев и не-больших речек. Высокое плодородие почв, поддерживаемое при-носом илистых частиц с прилегающих склонов долин и выходами обычно жестких грунтовых вод обеспечивают быстрый рост дре-весных пород: еловый древостой развивается по I классу бонитета. Именно здесь, в самых нижних частях склонов к долинкам ручь-ев, высота зрелых елей составляет 35 — 38 м. В то же время деревья не отличаются долговечностью, поскольку сформированная при быстрым росте рыхлая древесина в сочетании с высокой влажно-стью воздуха, очевидно, подвержена развитию гнилей, что при-водит к преждевременной гибели многих деревьев. Подлесок не характерен для приручьевых ельников, но при изреживании дре-востоя могут разрастаться крушина ломкая, жимолость, сморо-дина красная и черная, иногда подрост липы.
Травяной покров развит, главным образом, за счет крупных растений. Очень характерен страусник, местами образующий чи-стый покров. Распространены также борец, бодяки разнолистный и огородный, таволга, пролесник, кочедыжник женский крапива и другие виды. Деревья часто увиты хмелем. Во многих случаях заросли таволги настолько мощные, что некоторые исследовате-ли выделяют в качестве отдельного типа леса таволговый ельник (см. рис. 2.17).
Почвы — торфянисто-перегнойные, или торфяники, иногда с мощным слоем торфа, сильно оглеенные уже не небольшой глу-бине. Сильное переувлажнение почв препятствует освоению кор-
нями деревьев не только более глубоких слоев почвы, но в пони-женных местах и самого верхнего слоя, в результате чего создают-ся «мертвые» для корней деревьев зоны, занимающие большую или меньшую часть всей поверхности в зависимости от общего режима переувлажнения данного участка.

