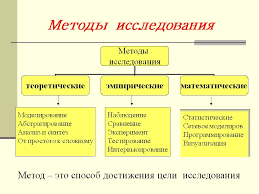
2015-04-06
2015-04-06 2011
20112.Цитокиндер, олардың түрлері және организмде қалыптасу принциптері. Интерферонды алу және жалпы сипаттамасы. Жоғарғы эукариотты ашытқылардың рекомбинатты цитокиндердің өнімі.
Өмірсүргіштігі төмен тимусқа тәуелді лимфоцитпен, макрофагтар және т.б. жасушалармен түзілетін биологиялық субстанцияларды цитокиндер (лимфокиндер) деп атайды. Цитокиндерге макрофагтардың тасымалдаушы және ингибридті мутациялық факторлар, интерлейкиндер, интерферондар және т.б. жатады.
Т-жасушаларымен және т.б. жасушалармен лимфокиндердің комплексі лимфоидты жүйеде аутоиммунды реакцияны және қарсы трансплантант реакциясын жүзеге асырады. Лимфокиндер жасушалық иммунитеттің медиаторлары болып табылады. Сенсибилизденген лимфоциттерді антигендермен ынталандырғанда, олар лимфокин деп аталатын (цитокин) еритін факторларды бөледі. Лимфокиндер үш топқа бөлінеді:
1Лимфокин- ингибитор, 2Лимфокин – стимулятор 4.Қабыну лимфокині
Цитоқиндер макрофагтармен байланыста болғанда сутегінің асқын тотығының жасушаларының өнімін ұлғайтып индукциялайды, бірақ бұл үрдісте каталаза ферментінің белсенділігі төмендейді.
Интерлейкин 1 мен 2-нің антиген немесе митогенмен бірігуі координацияланатын сигналдармен сипатталады, бұлардың лимфоидты жасушаның 3 типіне әсері бар, нәтижесі лимфоциттердің қуатты пролиферативті жауабы түрінде көрінеді. Цитокиндердің лимфоидты жүйе жасушасымен өзара әсері көміртегі, стероидты гормондар, фармакологиялық препараттар және олардың метаболитерімен тежелуі мүмкін. Цитокиндердің өнімдері гистосәйкестіктің басты қомплексімен бақыланады. Цитокиндер жергілікті әсер ету қасиетіне ие, ол кейде жалпы қан
ағымына түседі. Патогенді белгілері бар және антигеннің салмақтылығы басым болса, онда кейбір цитокиндер, мысалы, ИН-1, ИН-2, ісік некрозының факторлары қанға шығуы мүмкін. Жасанды жолмен организмге енгізсе, ол организмнен тез қайта шығады. Интерферонның қасиеттері.
Интерферон (ИФН) – ерекше вирусқа қарсы ақуыз түрі, олара зақымданған немесе тұтас организмнен өндіріледі. 1957 жылы ағылшын вирусологтары Айзикс пен Линденман ашқан. Бірнеше интерферон түрлері мен белоктардың класстары анықталған, олар бір-бірінен әртүрлі молекулярлық массалары арқылы ерекшеленеді. Вирустармен индукцияланған интерферонның молекулярлық массасы 26-38·10³ Д, ал полисахаридті бактериялармен индукцияланғандардікі 89-90·10³ Д болды. Антигендік телімділіктері бойынша интерферондар альфа (α), бета
(β), гаммаға (γ) бөлінеді. Интерферонның жасушада түзілу механизмі. Интерферон жүйесі телімді емес резистентіліктің маңызды факторы, ол организмнің барлық жасушасында көрсетілген және бөтен генетикалық ақпараты тану мен элиминацияға бағытталған. Интерферон индукциясы өлі және тірі вирустармен, табиғи нуклеин
қышқылдарымен, ситетикалық полирибонуклеидтармен, бактериялық антигендермен, кейбір полисахаридтермен және полианиондармен стимуляцияланады. Бұл процес ақуыз синтезінің ингибиторларына сезімтал болып келеді. Интерферогенез бірінен соң бірі жүретін 3 кезеңнен өтеді (индукция
-өнім-әсер) және дабылға жауап ретінде тізбекті реакциясын түзеді (дабылдар: инфекциялық, аллергиялық, аутоумунды, онкологиялық және т.б.) І фаза - интерферон индукциясы:
1. жасуша беткейіндегі индуктор адсорбциясы; 2. индуктордың жасушаны ұстауы; 3. индукцияның инициация процесі;
4. интерферон генінің дерепрессиясы; 5. интерферон үшін и-РНҚ-ның транскрипциясы (иРНҚ-ИФН).
ІІ фаза –интерферон өнімі: 1.транскрипция иРНҚ-ИФН; 2. трансляциялық 1-пост интерфероидтың түзілуі мен полипептидкеайналуы;4.интерфероидты гликолиздеу, интерферонның түзілуі; 5. интерферонның бөлінуі.
Қазіргі уақытта физиологиялық интерферондардың анықталуы мен зерттелуінің жан-жақтылығы, гомеостазды сақтауда бақылау мен реттеуге бағыттайды. ИФН-ы белгілі негізгі эффектісіне қарай антивирустық, антимикробтық, антипролифераттық, иммуномодульдеуші және радиопротективті деп бөлуге болады.
Вирустық материалдың генетикалық әр түрлігіне қарамастан, ИФН олардың кезеңдік репродукциясына басымдық көрсетеді, барлық вирустарға тән трансляцияның басын және вирусқа телімді ақуыздардың синтезін тежеп, шектейді. Осымен интерферонның антивирустық әсеріндегі ерекшелігі түсіндіріледі. Медициналық препараттардың ИНФ-ы құрамы бойынша альфа, бета, гамма болып бөлінеді, ал шығарылу уақыты мен қолданылуы бойынша – табиғи адамзаттың лейкоцитарлы интерферондар (бірінші ұрпақ ИФН) мен рекомбинантты болып бөлінеді (2-ұрпақ ИФН) Альфа және бета интерферондар белгілі антивирустың әсеріне, олар вирус ақуызының синтезін тежейді және репродукцияның басқа кезеңдеріне басымдылық көрсетеді. Гамма ИФН-дар иммуноциттерге белсенді әсер етеді (НК-жасушалар,
Т-лимфоциттер, моноциттер, монофагтар мен гранулоциттер), сондай-ақ гистосәйкестілік (НLА) комплексінің біріншілік және екіншілік антиген класстарының мембраналық эксрессиясын күшейтеді. ИФН көрсетілген әсер ету спектрін үлкен 3 топқа бөлуге болады: вирустық инфекция, ісік аурулары мен басқа да патологиялық формалары.
Иммунды және интерферон жүйелерінің ортасындағы байланыс айқындалды. ИФН фагоцитоз, табиғи киллердің белсенділігін, гистосәйкестіліктің басты комплексі антигеннің экспрессиясын стимуляциялайтыны көрсетілді. Екінші жағынан олар анафилаксиялық шоктың дамуын, қабынуды, гиперсезімталдылықтың баяу типі, антиденелердің түзілуін қыспаққа алады. Интерферондарды клиникалық қолдануда кері әсер етуі мүмкін:
тұмауға ұқсас белгілер, дерматологиялық реакциялар және гиперсезімталдылықтың белгілері және т.б. Сондай-ақ жеке органдармен жүйелерде де реакциялар тууы мүмкін: жүректің түтіктік жүйесінде, ішек-қарын жолында, зәр бөлетін жүйеде, орталық жүйке жүйесінде. Гендік инженерия әдістері шыққанша интерферондарды донорлық
қаннан алып келді, бір литр қаннан тазартылмаған 1 мкг интерферон алынады, ол бір инъекциялау дозасына жетеді.
Интерферонды вируспен түзілетін ауруларды емдеуге қолданылады, мысалы: герпес вирусымен, гепатит, сондай-ақ вирустарды инъекциялау жұмыстарында қолданылады. Олар сондай-ақ организмдегі ісік ауруларына (кеуде ісігі, тері, жұтқыншақ, өкпе және ми) емдік әсер етеді. Қазіргі кезде альфа, бета және гамма интерферондарын гендік инженериялық ішек таяшасын, жәндіктер мен сүт қоректілердің жасушаларының штамдарын қолданып сәтті алынуда.
3.Спирттік ашу. Спирттік ашудың қоздырушысы. Этил спиртін алудың технологиялық схемасы, ферментациялау шарттары және шикі зат көздері.
Спирттік ашыту. Өндірістік масштабта этанол химиялық, фармацевтикалық және тағамдық өнеркәсіптерді қолданылады., биоотын ретінде энергетикалық ресурс тұрғыдан да маңызды. Тағамдық саласында спирттік ашыту процессі шарап жасау, сыра дайындау, этанол өндіру, нан пісіру негізіне жатады. Спирттік ашу технологиясында ашытқының таза дақылы Saccharomyces cerevisiae, Saccharomyces vini, Saccharomyces cerevisiae var ellipsoids бактериялар Zimomonas mobilis сонымен қатар, Sacrina ventriculi, Erwinia amylovora және басқалар. Saccharomyces cerevisiae -нің төменгі және жоғарғы ашыту рассаларын ажыратады. Төменгі ажырату рассасына көптеген шарап және сыра ашытқылары жатады., ал жоғарғы ашыту рассассына – спирттік, наубайханалық және кейбір сыра ашытқылары жатады. Төменгі ашытқылар 6-100С активті, ал жоғарғылары 14-250С температураға бейімделген. Ашытқылар қышқыл ортада (рН 4-6) жақсы дамиды және активтігін 10-15% спирт ерітіндісінде сақтайды. Жоғарғы ашытқылар ашыту процессі кезінде жоғары мөлшерде көбік түзеді, СО2 бөлінеді, ашытқы массасы өзі көпірген сұйықтықтың үстіне шығады. Төменгі ашытқылардың қатысуы арқылы ашыту процессі бірқалыпты жүреді, ашытқы клеткасының массасы ыдыстың төменгі жағында қалады. Zimomonas spp. – грам оң таяқшалар, бұлар этанол концентрациясына төзімді, температуралық режимі 35-370С, алғашқы рет агава шырынынан дайындалған алгольдік сусын пулькадан бөлініп алынады, олар сонымен қатар пальма шырынында, қант қамысы шырынында табылған. Олар глюкозаны этанолға ашытқылардан да жылдам айналдырады, аздаған биомассасы бар, ашыту процессі кезінде аз мөлшерде сірке су және сүт қышқылдары пайда болады.
Klyveromyces flagilis ашытқыларын сүт сарысуындағы лактозаны ашыту арқылы спирт алу кезінде қолданылады. Глюкозаның этанолға айналуының 3 жолы белгілі: А – кетокластикалық (сүт қышқыл бактериялары), В – фосфорокластикалық (клостридалар, сарциналар), С – декарбоксикластикалық (ашытқылар)
Мұнда пируват аэробты тыныс алу сияқты ацетил СоА-ға айналмайды, бірақ пируватдекарбоксилаза көмегімен (негізгі фермент) ацетальдегидке дейін декарбоксилденеді. Ацетальдегид алкогольдегидрогеназа арқылы этанолға дейін тотықсыздандырылады.
Спирт түзілуінің кезеңдері:1.Алдын ала ферментацияға дайындау, шикізатты өндеу (субстрат). Өндірістік ашытқылар (бактериялар) штамдарының таза дақылын өсіріп-көбейтіп алу; 3. Биореакторда шикізаттың ферментациясы; 4.Этанолдың дистилляциясы.
1) Ашытқылар үшін көмірсутегінің негізгі алу көздеріне гексозалар, крахмал, инулин жатады: ал азот көздері – аммоний тұздары, мочевина, аминқышықылдар, ірі емес пептидтер, кейде – нитрат және нитриттер болады.
Ферментация процессінде қолданылатын шикізат үш типке бөлінеді: қантты, оны тура ашытуға болады (қант және крахмалдың өндірістегі қалдықтары – меласса, қант құрағы және қызылшаның шырыны);
- крахмалды, алдын ала өндеуді қажет етеді; қышқылдық немесе ферментативтік гидролизге ұшыратады (арпа, бидай суслосы, жүгері сиробы, күріш, қара бидай, сұлы). Ашытқыларда амилаза болмағандықтан ферментативтік гидролиз үшін бактериялық және мицелиалы амилаза пайдаланылады.
- қағаз-картон өнеркәсібінің қалдықтары, картоптық мезга, дәнді астықтардың сабаны, жүгері собығы.
2) Периодтық ферментация 48-72 сағат жүреді.. ферментация процессінің басында алдымен ашытқы биомассасын жинау үшін аэробты жағдай жасайды, осыдан кейін аэрацияны тоқтатады..
Оттегі кірген соң ашытқылар көмірсуды тотықтыра бастайды, яғни ашыту процесінен аэробты тыныс алу процесіне ауысады. Аэробты жағдайда пирожүзім қышқылынан ацетил СоА, үшкарбон қышқылдар циклы арқылы субстраттың СО2 мен Н2 О дейін толық тотығуы жүреді, бұл ашытқылар үшін энергетикалық тиімді, олар жақсы өседі, биомасса көп жиналады; үш карбон қышқылдар циклі барысында клетка синтезі үшін қажетті метаболиттер көп түзіледі.
Анаэробты жағдайға ауыстырғанда этанол 3-4 есе артады, өйткені тыныс алу мен субстратты фосфолирленуі арасында еркін АТР талас тоқтайды; гликолиз жолына энергия мен шикізаттың көп мөлшері қажет.
Ашыту процессі аяқталған уақытта этил спирті 14-16% мөлшерде жиналып, ашытқылардың әрі қарай көбеюін тежейді.
3) Дистилляция – ұшпа компоненттердің сұйық және қатты бөлшектерден ажыратылуы, дистилятта этанолдың салмағы концентрлеу және ректификация бойынша 30-96% дейін болады.
Этанолдың суға қарағанда қайнау температурасы төмен, буда этанол концентрациясы суға қарағанда жоғары. Мысалы, егер де су ерітіндісінде этанол 10% болса, будағы салмағы – 50% тен, яғни дистиллят этанолмен қатты байытылған. Максималдық дистилляция 78,20С температурада болады және салмағы 96% тең, жоғары болмайды.
4.Ағынды суларды биотехнологиялық тазалау. Жер асты және жер үсті суларының ластану түрлері. Ағынды суларды тазалау әдістері. Ағыс жолдарын тазалаудың аэробтық жүйесі. Ағынды суларды тазартудың кезеңдік үрдістері - Алдын – ала механикалық тазарту – тор арқылы сүзу, ірі бөлшектерін тұндыру, майлар флотациясы. Механикалық өңдеу нәтижесінде ағынды сулардан жалпы ластану массасының 30% – ға жуығы жойылады; - Биологиялық тазарту – алдымен активті лайдың аэробты және анэробты микроорганизмдермен ластанудың ыдырауы; - Химиялық тазарту – ағынды сулардан фосфаттарды темір мен алюминий (FePO46 AlPO4) тұздарының көмегімен алып тастау. Фосфаттардың бір бөлігің АТР түзілу энергия қоры қызметін атқаратын клетка ішілік полифосфат түрінде анаэробты (аценетобактериялар және басқалар) жинайды.
Жалпы тазарту орталықтары механикалық тазарту (тор, озондау) технологиясы қолданылады.
Биологиялық тазартудың негізгі міндеттері органикалық көміртегін жою, аммияқ тотығуы және ағын сулар қабылдайтын табиғи суда оттегі шығынының алдын – алу үшін еріген, байланысқан азотты жою. Органикалық ластанулардың ыдырау үрдісінде түзілетін аммиак, мочевиналар нитрификациялайтын бактериялармен нитрит және нитраттарға дейін тотығады. Кейін анаэробты жағдайда нитраттарға азотқа дейін тотықсызданады және іркінді сулардан жойылады.
Биологиялық тазарту үрдісі екі кезеңнен тұрады: тұңған ағынды судың ауа оттегісімен және аэротенкада (бірнеше сағаттан бір тәулікке дейін) активті лай микроорганизмдер өзара әрекеті;тазартылған, түсіздендірілген ағынды суды ауасыздандырылмаған тұндырғышта (бірнеше тәуліктен біренше аптаға дейін) белсенді лайдың бөлшектерінен бөлу, белсенді лайдың тығыздануы. Активті лайдың бір бөлігі келіп қосылатын ағын суда органикалық ластаудың әсері ыдырау үін аэротенкте тұңбадан әрдайым қайтып оралады (рециклизация).
Ағынды суларды тазалаудың биологиялық әдісі Ағынды сулар құрамына кіретін әр түрлі қосылыстарды субстраттар ретінде микроорганизмдердің қолдану қабілетіне негізделген әдіс.
Бұл әдістің артықшылығы: -Органикалық және бейорганикалық заттарды жою мүмкіндігі-Аппараттар мен өту үрдістің қарапайымдылығы-Ағынды суларды биологиялық жолмен тазартудың 2 үрдісін қолданылады:
- Аэробты - мұнда микроорганизмдер заттарды тотықтыру үшін оттегіні қолданылады. Бұл үрдіс екіншісіне қарағанда сенімді. - Анаэробты - бұл кезде микроорганизмдер бос ерітілген оттегіне қол жеткізе алмайды. Электрон акцепторы ретінде олар органикалық заттардың көміртегін қолдана алады. Бұл үрдіс баяу өтетіндіктен, өте қымбат және көлемді құрылғыларды қажет етеді. Ағынды суларды тазартудың аэробты үрдісі
Тазартудың аэробты үрдісінде микроорганизмдермен тотыққанда органикалық заттардың бір бөлігі биосинтез үрдісінде қолданылады, басқасы – қауіпсіз өнімге айналады – H2O, CO2, NO2 және тағы басқаларға. Стоктардың биологиялық тазартылуы конструкциялары әр текті құрылғыларды жүргізіледі – биофильтрлер мен аэротентер. Биофильтрлер – биологиялық тазарту аппараты болып табылады. Биофильтрлер арқылы тазарту:
1. Биопленкамен тіркестіру;
2. Микроағзалардың жасушалардың үстіне органикалық заттардың сорбциясы;
3. Микроағзалардың метоболизміне байланысты ағынды сулардың тотықтануы.
Ағынды сулардан металдарды айыратын бактериалар
Микроорганизмдердің кейбір түрлері сұйықтықта ерітілген металдарды сорбциялай алады, металдардың концентрациясы жоғары болады, ал жылулық өндеуден кейін биосорбты металдарды қолдануға мүмкіндік болды. Ағын суларды тазартуда спиртті май қышқылдары, парафинді, ароматты көмірсуларды, фенолды тотықтыратын аэробты және анаэробты микроорганизмдерді қолданады. Соның ішінде Actinomyces, Actinobacter. Bacillus. Pseudomanos. Corinobacterium т.б.
5.Сомаклоналды өзгергіштік, сомоклоналды варианттар. Сомоклоналды варианттарды селекциялық үрдісте қолдану.
Сомаклондық өзгергіштік - өсімдік жасушаларыныѕ ядролық және органоидтық геномдарыныѕ тұрақсыздығынан туындайтын фенотиптік өзгерістер. In vitro жағдайында өсімдіктіѕ қалпына келтіру әдістерін дамыту және сомаклоналдық өзгергіштік құбылысыныѕ ашылуы қоздырылған мутанттарды алуға мүмкіндік берді.
Сомаклоналдық өзгергіштік ұлпаны қолдан өсіргенде пайда болатын өзгергіштіктіѕ бірі. In vitro өсіндісінде алынған ұлпаныѕ бірқалыпты болмауы туралы алғашқы мәліметтер 1961 жылы жарияланған. Кейбір жағдайда қалпына келтірілген өсімдіктіѕ жеке қасиеттері жағынан бастапқы өсімдіктен 15-20% айырмашылығы болады.
Сомаклоналдық өзгергіштіктіѕ ерекшелігі Сомаклоналдық өзгергіштік кариотиптік, морфологиялық, биохимиялық және молекулалық деѕгейлерде байқалады. In vitro жасушаларында полиплоидтық және анеуплоидтық өзгерістер, хромосомалық аберрациялар, митоздыѕ бұзылуы белгілі болған. Жеке диплоидтық жасушалардан алынған каллус ұлпасында диплоидты, тетраплоидты және анеуплоидты хромосомалар жиынтығы анықталған. Хромосомалардыѕ үлкен және кіші өзгерістері анықталған кезде, транслокация және анафаза қөпіршелері пайда болып, хромосомалар ажырайды. Хромосомаларды дифференциалдық тәсілмен бояғанда, оларға тән құрылымдық өзгерістер айқындалды.
Генетикалық өзгерістер деѕгейін көтеру және жаѕа гендердіѕ пайда болу мүмкіндігініѕ биологиялық маѕызы зор, өйткені бұл өсімдік селекциясында жаѕа зерттеулерді жүргізуге жол ашады. Сомаклоналдық өзергіштік арқылы генқорды байытуға мүмкіндік туады. Сонымен бірге сомаклоналдық өзгергіштіктіѕ көмегімен, селекциялық жолмен бір генотипте қосылмайтын белгілерді алуға болады, мысалы күріштіѕ тез піскіштігі мен ұзын дәнділігін немесе күріштіѕ тез піскіштігі мен төмен температураға төзімділігін біріктіретін түрді алуға болады. Қоздырылған мутагенез сияқты сомаклоналдық өзгергіштік те белгілі бір бағытта болмайды. Дегенмен, қалпына келтіру кезінде зиянды өзгерістердіѕ біразы жойылып кетеді. Сомаклоналдық өзгергіштіктіѕ арасында күрделі де, қарапайым генетикалық өзгерістер кездеседі. Мәселен, картоп бұтағыныѕ пішінін, түйнек бояуын және оныѕ түйін байлау мерзімін, ауруға төзімділігін, фотокезеѕге сұранысын өзгертетін; темекі мен қызанақтыѕ пішінін, гүл, жеміс және жапырақ бояуын, пісу мерзімін, аталықтыѕ ұрықсыздығын өзгертетін сомаклондар алынды. Жұмсақ бидай регенеранттары арасында тұқым қуалайтын мынадай белгілерінде өзгерістер байқалған: масақ биіктігі, масақтағы дәндер саны, ақуыз бен ферменттердіѕ қаѕықпасы. Гендік талдау көрсеткендей, сомаклоналдық өзгергіштіктіѕ генетикалық негізі көптеген жағдайда ядро, пластид немесе митохондрия геномындағы бірліөжарым гендердіѕ немесе олардыѕ шағын тобыныѕ доминант, жартылай доминант немесе рецессивті мутациялары болып табылады.
Жасуша өсіндісінде фенотиптік өзгергіштіктіѕ пайда болуы өсуді реттейтін механизмніѕ бұзылуымен түсіндіріледі. Кейбір осындай өзгерістер митоз арқылы ұрпақтан ұрпаққа берілуі мүмкін, бірақ қалпына келтірілген өсімдіктерде немесе жынысты жолмен көбейгенде бұл өзгерістер байқалмайды. Гаметадан түзілген өсімдіктерде генетикалық өзгергіштік пайда болады, оны гаметоклоналдық өзгергіштік деп атайды. Гаметоклоналдық өзгергіштік құбылысы және оныѕ селекциядағы маѕызы тозаѕқап өсіндісі деген арнаулы тақырыпта кеѕінен қарастырылады. Сомаклоналдық өзгергіштікті туғызатын факторлар Жасушаны өсірген кезде оныѕ өзгергіштік ауқымына өсімдіктіѕ генотипі қатты әсер етеді. Сомаклоналдық өзгергіштік көптеген жағдайда генетикалық конституцияға байланысты екені белгілі.
Сомаклоналдық өзгергіштік деѕгейі қоректік ортаға отырғызылатын эксплант алғашкы өсімдіктіѕ қай мүшесінен: өркен төбесі, жапырақтыѕ астыѕғы жағы, сағақ, сабақ буыны, гүл бүршігі, жетілмеген бүршік, жарнақтар, тозаѕқаптан алынғандығына байланысты. Меристема өскінінде бұл құбылыс аз байқалады, ал сұйық ортада өсірілетін протопласттарда немесе маманданбаған каллус ұлпасында көбірек кездеседі. Мутагенезді қоздыруда жасанды қоректік ортаныѕ, әсіресе фитогормондардыѕ маѕызы зор. Фитогормондар жасушаныѕ өсу айналымына ықпал жасап, митоз үрдісін өзгертеді. Тұтас өсімдіктіѕ өсу жағдайлары мен бағдарламасын бұзып, оныѕ бір мүшесін жасанды коректік ортаға отырғызу сомаклоналдық өзгергіштікті туғызады. Ұлпа мен жасушаны іn vitro жағдайында көп уақыт өсіру де сомаклоналдық өзгергіштік деѕгейіне әсер етеді. Мысалы сұлы регенеранттары арасында өсіндініѕ мерзімініѕ ұзаруы әсер еткен өзгерістер кездеседі.
Өсімдік селекциясында сомаклоналдық өзгергіштікті пайдалану Сомаклоналдық өзгергіштіктіѕ артықшылығыөөсімдіктіѕ бағалы сорттарында будандастырусыз қосымша генетикалық бағалы өзгергіштіктіѕ пайда болуында.Сомаклоналдық өзгергіштікті пайдаланудыѕ екі жолы бар. Біріншісі – in vitro жағдайында селекциясыз сомаклоналдық өзгергіштікті сұрыптау. Сомаклоналдық өзгергіштікті туғызатын себептерге байланысты регенерант өсімдіктерінде өзінен өзі пайда болатын кездейсоқ өзгерістер байқалады. Бұл өзгерістерді бағалы белгілері бойынша сұрыптап отырып, қалыптасқан селекция үрдісіне енгізеді. Осыныѕ нәтижесінде бағалы сорттарды алады. Екіншісі – in vitro селекциясыныѕ көмегімен, яғни жасуша селекциясы арқылы сомаклондарды сұрыптау. Жасуша селекциясы - in vitro өсіндісінде бір түрдегі жасушалардыѕ көбеюіне байланысты бағалы белгілерді сұрыптау. Егер осындай жасушалар қолайсыз жағдайға төзімді болса, сол жасушалардан пайда болған регенерант өсімдіктері де төзімді болады. Басқа әдістер сияқты жасуша селекциясыныѕ да кемшіліктері мен артықшылықтары бар. Мысалы, қалыптасқан селекциямен салыстырғанда, Петри тостағанындағы жасуша популяциясын жеке ағза деп санасақ, бір тәжірибеде миллиондаған ағзамен жұмыс жүргізуге болады, ал қалыптасқан селекцияда селекционер қолайлы жағдайда тек мыѕ өсімдікпен жұмыс жасайды. Жасуша селекциясыныѕ өте құнды артықшылығыныѕ бірі – қалыптасқан селекция әдісімен сұрыпталмайтын пайдалы белгілерді алуға болады. Жасуша селекциясыныѕ кемшіліктерініѕ бірі – жасушаныѕ хромосомалық және молекулалық деѕгейіндегі зертеулердіѕ өзара байланысы туралы мәліметтердіѕ мардымсыздығы. Екіншіден, жасуша селекциясы әдісін пайдалану үшін, мысалы, топырақ тұздылығына төзімді өсімдік алу кезінде оны тудыратын механизмдер жасуша және тұтас өсімдік деѕгейінде де кездесу керек. Себебі жасуша селекциясында мынадай жағдай жиі кездеседі, мысалы, жасуша деѕгейінде төзімділік байқалады, ал оны тұтас өсімдік деѕгейіне көшіргенде бұл пайдалы белгі жойылып кетеді. Үшіншіден, жасушаны in vitro өсіндісінде ұзақ уақыт өсірсе, оныѕ морфогендік, яғни жасушаныѕ тұтас өсімдікті қалыптастыру қасиеті жойылады. Мысалы, бидай өсіндісінде 8-9-шы отырғызудан кейін каллус ұлпасыныѕ морфогендік қасиеті жойылып кетеді. Төртінші кемшілігі жасушалар селективтік факторға үйреншікті болып, қарапайым төзімділікті туғызуы мүмкін.