 2015-05-06
2015-05-06 870
870У большинства животных половые железы имеют весьма яркое формообразовательное значение. Развивающиеся под влиянием половых гормонов вторичные половые признаки особенно выразительны у многих птиц, а нередко и у млекопитающих. Во многих случаях самцы и самки сильно отличаются по своей внешности — по общей величине и форме тела, окраске перьев или шерсти, различным придаткам и выростам (гребни, шпоры, перья у птиц; рога, бивни, клыки у млекопитающих), голосу, поведению и т. п.
Опыты кастрации птиц, а также эксперименты с пересадкой половых желез показали, что формы зависимости развития вторично-половых признаков бывают различными. Так, например, У Домашних кур головные придатки петуха (гребень, бородки и сережки), его пение и поведение развиваются под влиянием мужского полового гормона (зависимые половые признаки по М. М. Завадовскому), а яркое оперение петуха и его шпоры развиваются и без влияния гормона (независимые половые признаки). С другой стороны, женский половой гормон подавляет раз-витие мужских независимых признаков, т. е. шпор и петушьего °перения, и стимулирует развитие яйцевода, типичного головного Уоора и оперения курицы. Вместо пересадки желез можно производить инъекции вытяжки этих желез.
Половые гормоны лишены видовой специфичности. Они, например, с успехом могут быть заменены гормонами млекопитающих (включая человека) или синтетическими гормонами (тесто-
290 Регуляция формообразования в индивидуальном развитии
стероном и эстроном). Однако действие их строго специфично, как по месту и форме реакции (что зависит от дифференцировкц реагирующей ткани), так и по видовой их специфике. При любых пересадках и комбинациях под влиянием мужского полового гормона (например, человека) в трансплантате развиваются мун^ ские половые признаки (например, оперение) того вида и даже той породы или мутации, от которой взят трансплантат.
Такие же эксперименты проводились и над другими позвоночными, начиная с рыб и кончая млекопитающими. В случае выраженного полового диморфизма кастрированные самцы млекопитающих приобретают наряд самки. С другой стороны, и у самки под влиянием мужского полового гормона могут развиться характерные самцовые признаки. (Подобные же отношения описаны и у некоторых беспозвоночных — червей, ракообразных. Между тем у насекомых подобный же половой диморфизм реализуется независимо от половых желез — кастрация и пересадка желез не ведут к изменению внешних половых отличий.)
Дальнейшие исследования показали, что в отношении половых гормонов нет качественных различий между полами. Как яичники, так и семенники продуцируют оба половых гормона, но только в разной концентрации. В результате у самки преобладает женский гормон, а у самца мужской. Признаки того или другого пола развиваются только под влиянием некоторого, необходимого для этого минимума концентрации гормонов, когда достигается пороговый уровень нормальной реакции данной ткани.
Это дает возможность частичного извращения половых различий и объясняет нередкие случаи переноса половых признаков с одного пола на другой. У амфибий половые железы развиваются вначале как обоеполые железы, и у них действием половых гормонов можно вызвать полное преобразование пола. У лягушек и квакш действием тестостерона самки могут быть превращены в настоящих самцов, которые при скрещивании с самками дают, однако, исключительно самок. Однако примечательно, что тот же результат может быть достигнут применением высоких концентраций женского гормона — эстрона. Наоборот, у тритонов действием эстрона самцы превращаются в самок, которые при оплодотворении самцами дают исключительно самцов. Эти реакции определяются наследственной структурой клеточного ядра (XX у самок лягушек и ZZ у самцов тритонов). У других амфибии в подобных же экспериментах получается лишь частичная маскулинизация самок и феминизация самцов.
У оленей рога являются характерным половым признаком зрелого самца. Однако у северного оленя рога имеются и у самки. Это, несомненно, их ыовоприобретение — результат вторичного переноса самцового признака на самку. В Саянах сохранилась даже исходная форма северного оленя, у которой самки безроги
Глава IV. Гормональная регуляция роста и формообразования 291
/Мангковцев, устное сообщение). Рога оленей всегда развиваются д влиянием мужского полового гормона, который должен достигнуть ко времени полового созревания известной пороговой концентрации (рис. 23). Мужской половой гормон выделяется и Y самки, однако не достигает концентрации, характерной для самцов. У северного оленя имеется то же самое, однако у него изменилась норма реакции тканей на мужской половой гормон.
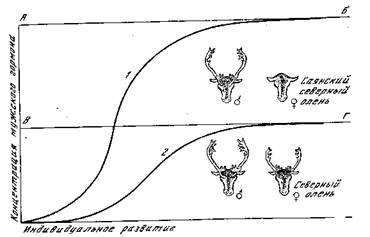
Рис. 23. Развитие рогов у северного оленя [Машковцев]
АБ — пороговый уровень реакции у саянского оленя; ВГ — пороговый уровень реакции у обычного северного оленя; 1 — кривая нарастания концентрации мужского гормона у самца; 2 — то же, у самки
Морфогенная реакция, выражающаяся в образовании рогов, наступает у них на более низкой концентрации гормона вследствие снижения порогового уровня реактивности тканей. Концентрация мужского полового гормона самки оказывается достаточной для развития типичного самцового признака — рогов. Пример формообразовательного действия половых гормонов показывает нам вновь возможность возникновения качественных преобразований на основе чисто количественных различий и значение пороговых уровней для осуществления известных реакций. Железы внутренней секреции изучены у человека и у млекопитающих гораздо лучше, чем у других животных, и показывают аам исключительно сложную картину их взаимодействия, охватывающего как стимуляцию, так и торможение секреторной Функции. Эндокринные регуляции находятся также под контролем нервной системы и сочетаются с влиянием внешних факторов. Если ограничиваться лишь рассмотрением морфогенного влияния половых желез, то функция последних контролируется
292 Регуляция формообразования в индивидуальном развитии
прежде всего деятельностью передней доли гипофиза. Именно гормон передней доли гипофиза стимулирует половое созревание и определяет периодичность функции половых желез. Функция гипофиза, со своей стороны, контролируется нервной системой, а через посредство органов чувств также и условиями внешней среды. Среди факторов последней, определяющих половую деятельность, особое значение имеют температура, длительность светового дня, а также непосредственные зрительные, звуковые восприятия и запахи, доносящиеся от особей другого пола. Как видно, система взаимозависимостей, определяющая формообразование, а в том числе и процессы полового созревания и развития вторичных половых признаков, в высшей степени сложна. Для разбираемых нами вопросов важно отметить, что по меньшей мере через посредство нервной системы осуществляется и обратная связь, т. е. контроль половых реакций, за которым следует их регуляция со стороны гипофиза.
Обратная связь осуществляется иногда и по гуморальным путям от вторичных половых признаков. Так, например, у петухов, как мы знаем, гребень развивается под влиянием полового гормона, выделяемого семенниками. Однако на известной стадии развития гребень оказывает уже задерживающее влияние на развитие семенников. Задержка в развитии семенников вызывает прекращение роста гребня. Если удалить половые железы — редуцируется гребень. Если же удалить гребень — усилится рост желез. В данном случае, следовательно, имеется отрицательная обратная связь от гребня к половым железам. Такая форма связи характерна для зрелого организма и ведет к поддержанию подвижного равновесия в системе, достигшей стационарного состояния. Вероятно, таковы же соотношения и при развитии рогов у оленей. Молодые, растущие рога оленей (панты) являются, несомненно, органами внутренней секреции, гормоны которых, по-видимому, служат средством контроля их роста и развития, осуществляемого с помощью секреторной функции половых желез (вероятно, через посредство гипофиза).
На примерах действия некоторых гормонов у животных мы установили вновь наличие тех же закономерностей, которые уже отмечались нами при разборе форм взаимодействия частей в развитии зародыша и при рассмотрении гормональных регуляций У растений: 1) отсутствие видовой специфичности индуцирующих веществ (гормона или индуктора) и широкую их заменяемость рядом химически сходных веществ; 2) наличие органотипичной специфики реакции, зависящей от дифференцировки реагирую' щей ткани (гребень, бородка), и 3) строгую видовую специфичность результата реакции, определяемую унаследованной ее нормой, характерной для вида, породы, мутации.
Следует еще отметить явную зависимость всей системы гормональной регуляции от внешних факторов, дающей известную
"W-
Глава V. Взаимодействие между ядром и цитоплазмой 293
настройку всему организму в целом (на температуру, пищу, свет и т. д.)- Это ясно в отношении гипофиза, щитовидной и половых желез. При малой специфичности индуцирующих веществ и гормонов естественно, что факторами, определяющими специфику морфогенетических реакций, являются, с одной стороны, специфические формы обмена в дифференцированных тканях (органо-типическая специфика реакции) и, с другой стороны, специфика клеточного обмена, контролируемая унаследованными субстанциями ядра и цитоплазмы (наследственная норма реакции).
Глава V
ВЗАИМОДЕЙСТВИЕ
МЕЖДУ ЯДРОМ И ЦИТОПЛАЗМОЙ
КАК ОСНОВА СПЕЦИФИЧНОСТИ
ФОРМООБРАЗОВАТЕЛЬНЫХ РЕАКЦИЙ
Весь рассмотренный материал показывает нам:
1) зависимость структуры яйца или зачатка от факторов внешней среды, питающих данное яйцо или зачаток;
2) зависимость формообразования от строения исходного материала — яйца или зачатка, имеющего по меньшей мере полярную структуру с соответствующим распределением некоторых более или менее активных веществ;
3) значение формообразовательных движений, приводящих к новым соотношениям с внешней средой, к новым взаимосвязям между частями самого зародыша или зачатка;
4) зависимость формообразования от положения и взаимодействия различных частей, при котором происходит передача активных веществ цитоплазмы от одной части к другой (индукция), а также обмен продуктами метаболизма;
5) значение обратных связей между развивающимся зачатком и другими частями органа или зародыша, определяющими установление нормальных соотношений между ними. Эти связи могут проявляться во взаимной стимуляции, ведущей к прогрессивному развитию всей системы. По мере достижения известного уровня развития такое взаимодействие прекращается или заменяется обратной связью, когда одна часть оказывает на другую тормозящее влияние;
6) малую специфичность веществ, оказывающих стимулирующее или тормозящее влияние на развитие. Эти формообразующие вещества лишены видовой специфичности, т. е. могут быть заменены аналогичными веществами других, подчас очень далеких,
294 Регуляция формообразования в индивидуальном развитии
организмов. Из индуцирующих веществ наибольшее значение имеют белки и нуклеопротеиды. В случае гормонов это относительно простые химические соединения, которые могут быть заменены другими сходными соединениями. С другой стороны, нельзя говорить о полной неспецифичности таких веществ — в каждом случае типичная реакция вызывается только определенной категорией химических субстанций. По-видимому, некоторые реакции требуют для своего осуществления наличия определенных нуклеопротеидов;
7) специфика реакции, если она только может реализоваться (т. е. если имеются требуемые для этого условия, как внешние, так и внутренние, включая наличие индукторов и т. п.), определяется, однако, исключительно реагирующим материалом. Эти реакции обнаруживают, во-первых, видовую, или, точнее, геноти-пическую, специфику и, во-вторых, органотипическую специфику, различающуюся в разных тканях и органах.
Видовая, или, точнее, генотипическая специфичность реакции (касаясь не только видовых, но и расовых и мутантных признаков) определяется (контролируется) генотипом. Органотипиче-ская специфика определяется взаимодействием частей на основе уже существующих дифференцировок.
В общей форме мы можем сказать, что все формообразование покоится на унаследованной структуре яйца, на ее взаимодействии с внешней средой и на взаимодействии различных веществ яйца и частей развивающегося зародыша. Специфика всех процессов развития определяется, с одной стороны, положением и дифференцировкой данной части, ведущими к органотипическому выражению формообразовательных реакций, и, с другой стороны, унаследованной нормой реакций, ведущей к реализации видовых, расовых и мутантных признаков организма. Такое противоположение генетической и органотипической специфики формообразования вполне закономерно с точки зрения теории развития. Так, например, видовые особенности оперения птицы определяются, несомненно, ее генотипом (и весьма отличны не только у разных видов птиц, но и у разных мутаций и разных пород, например, домашних уток, гусей и кур). Однако это оперение реализуется по-разному в различных частях тела, образуя характерный покров на спине, брюхе, голове, выделяя особые формы пуховых и контурных перьев на туловище, покровных и маховых перьев разного порядка на передних конечностях и рулевых— на хвосте. Вся эта различная, реализация видовых признаков определяется местными условиями развития зачатков перьев в различных органах. Различие в развитии и в конечном результате совершенно ясное. Вместе с тем, однако, нельзя разделять и отрывать эти явления друг от друга — видовые признаки познаются нами только в их органотипической реализации, которая (как и вся организация) является необходимой частью ха-
Глава V. Взаимодействие между ядром и цитоплазмой 295
рактеристики вида. С другой стороны, в любом органе каждая структура является выражением не только местных условий развития, но и генотипических особенностей данного вида, расы, мутации.
Мы можем сказать, как это обычно делается, что развитие генотипических особенностей зависит от унаследованной нормы реакции, т. е. от ядерных структур. Вместе с тем, мы вправе сказать, что реализация всех органотипических и гистологических структур зависит от плазматических субстанций яйца, их распределения, от специфики обмена веществ. Однако и в этом обычном противопоставлении значения ядерных структур и специфики цитоплазматических субстанций, определяющих клеточный обмен веществ, мы ни в коем случае не можем отрывать их друг от друга. Ядерные вещества могут проявлять свою деятельность только в связи с цитоплазмой, а формообразовательные функции цитоплазмы, несомненно, контролируются ядерными структурами. Ядро и цитоплазма только совместно определяют специфику формообразования. Поэтому нам надлежит теперь ознакомиться более обстоятельно с формами взаимодействия между ядром и цитоплазмой в процессах детерминации и формообразования.
Для исследования этих вопросов производились многочисленные эксперименты удаления ядра из клеточного тела и пересадки ядра из одной клетки в другую. В первом случае изучались формообразовательные потенции изолированной цитоплазмы, во втором случае изучались результаты совмещения одинаковых ядер с различными цитоплазмами или разных ядер с одинаковыми цитоплазмами. Удаление ядра может производиться путем его инактивации действием рентгеновых или ультрафиолетовых лучей или, на больших объектах, путем его разрушения или даже путем непосредственного извлечения микропипеткой. Совмещение ядра с различными цитоплазмами возможно путем оплодотворения яйца с удаленным или уничтоженным ядром или путем непосредственной пересадки микропипеткой ядра в клетку, предварительно лишенную своего собственного ядра.
Замечательным объектом для экспериментального изучения значения ядра и цитоплазмы для формообразования оказались крупные одноклеточные водоросли рода Acetabularia (рис. 24). Эти водоросли обладают сложно дифференцированным телом, состоящим из длинного стебля с ризоидами на нижнем конце и сложно расчлененной шляпкой на верхнем. Из зиготы Acetabularia mediterranea развивается проросток, образующий стебелек и ризоиды, которыми он прикрепляется к субстрату. По мере Роста стебелька в длину на его свободном конце развиваются мутовки разветвленных волосовидных окончаний. Мутовки недолговечны — они быстро отмирают, а на их месте последовательно развиваются все новые мутовки волосков до тех пор7 покд
296 Регуляция формообразования в индивидуальном развитии
продолжается рост стебля. Когда рост стебля закончен, под мутовкой волосков закладывается круг радиально расположенных гаметангиев, которые все вместе образуют шляпку. Когда шляпка достигла нормального размера, ядро повторно делится и образует тысячи вторичных ядер. После этого возникает восходящий ток плазмы, возносящий ядра и хроматофоры, которые почти все распределяются в камерах шляпки. В камерах образуются многие (около 15 000) цисты. После многократного деления ядра каждой цисты образуются многочисленные гаметы (около 1800 гамет в цисте). Различные виды Acetabularia различаются, между прочим, по величине и строению шляпки. Они удивительно хорошо регенерируют и допускают разнообразные опыты сращения частей как в пределах вида, так и между разными видами.

Рис. 24. Развитие Acetabularia mediterranea [Кюн, 1955] l—ll — стадии развития
Глава V. Взаимодействие между ядром и цитоплазмой UQ7
Регенерируют даже безъядерные отрезки. В них возможен настоящий рост, связанный с синтезом белков и размножением хроматофоров. Возможно и формообразование. Возможно образование шляпки на апикальном конце и образование ризоидов у основания стебелька. Рядом последовательных опытов доказано, что образование шляпок или ризоидов зависит от распределения вдоль стебля специфических формообразующих веществ, которые образуют два противоположных градиента (рис. 25). От уровня градиента зависит образование либо шляпки, либо ризоидов (промежуточные органы в этом случае никогда не образуются).
Если срезать уже образовавшуюся шляпку, то деление ядер в ризоидах прекращается, регенерирует шляпка, и только после ее полного развития возобновляется деление ядер, а вслед за тем возникает и восходящий ток цитоплазмы. Таким образом, деление ядер, а также движение цитоплазмы зависят от завершения формообразования шляпки. От готовой шляпки к ядру, очевидно, имеется обратная связь, служащая сигналом для начала деления.
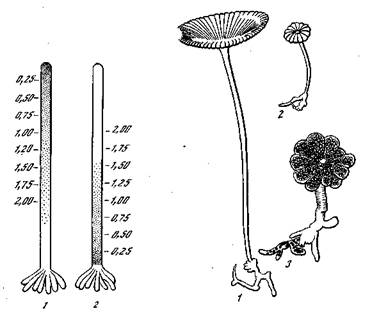
Рис. 25. Схема распределения концентрации формообразующих веществ у Acetabularia [Кюн, 1955] 1 ~~ Для шляпок; 2 — для ризоидов Рис. 26. Пересадки у Acetabularia [Кюн, 1955]
1 — Acetabularia mediterranea, X 5; 2 — Acetabularia wettsteinii X 5; 3 — безъядерный стебелек A. mediterranea, пересаженный на ядросодержащий ризоид A. wettsteinii, х 12
298 Регуляция формообразования в индивидуальном развитии
Опытами двойной регенерации и пересадок доказано, что формообразующие вещества синтезируются под влиянием ядра (рис. 26). Трансплантации между Acetabularia mediterranea и Acetabularia wettsteinii показывают, что эти вещества обладают видовой специфичностью формообразования, т. е. в плазме Acetabularia mediterranea ядро Acetabularia wettsteinii выделяет вещества, образующие типичную шляпку Acetabularia wettsteinii. Возмояшо и образование смешанных форм шляпок при соединении больших стеблей Acetabularia mediterranea с ядром Acetabularia wettsteinii, так как в этих стеблях был уже достаточный запас формообразующих веществ Acetabularia mediterranea. Однако при удалении такой промея^уточной шляпки регенерирующая новая шляпка образуется полностью по типу Acetabularia wettsteinii, соответственно принадлежности ядра. При двухъ-ядерных комбинациях между разными видами Acetabularia (одно ядро Acetabularia mediterranea+одно ядро Acetabularia crenula-ta) получаются промежуточные шляпки. При трехъядерных комбинациях (два ядра Acetabularia crenulata и одно ядро Acetabularia mediterranea) развиваются шляпки, более сходные с Acetabularia crenulata. Во всех этих экспериментах выявляется значение ядра, как носителя видовых свойств, которые реализуются, однако, через посредство формообразовательных веществ цитоплазмы. Можно утверждать, что связь между ядром и цитоплазмой осуществляется в этом случае при самом активном участии специфических рибонуклеиновых кислот. В пользу этого говорят следующие факты: трипафлавин, который прочно связывается с нуклеиновыми кислотами, подавляет регенерацию как безъядерных, так и содержащих ядро частей Acetabularia.
Ультрафиолетовые лучи (2537 А) сильно поглощаются нуклеиновыми кислотами и в то же время подавляют регенерацию Acetabularia. Рентгеновы лучи, которые разрушают ДНК, но сравнительно мало действуют на РНК, мало влияют и на регенерацию безъядерных частей. Рибонуклеиновые кислоты выступают здесь либо прямо, либо косвенно (через синтез специфических белков) в роли формообразующих веществ, обладающих видовой специфичностью. Образование рибонуклеиновых кислот и их специфичность определяются, однако, ядром. Аналогичные эксперименты со сходными результатами производились и над другими водорослями и над простейшими животными.
Взаимодействие между ядром и цитоплазмой имеет, следовательно, характер замкнутого цикла взаимозависимостей: ядро определяет синтез РНК с видовой специфичностью, которая либо прямо, либо через синтез специфических белков определяет формообразование (шляпки и ризоида). Формообразование шляпки оказывает обратное влияние на ядро, вызывая его последовательное деление и последующие движения цитоплазмы, переносящие ядра в камеры шляпки.
Глава V. Взаимодействие между ядром и цитоплазмой 299
Вернемся теперь вновь к организации яиц животных и рассмотрим вопрос о формообразовательном значении их ядра и цитоплазмы. Одним из самых замечательных примеров взаимодействия ядра и цитоплазмы в процессах развития является изученное Бовери еще полстолетия тому назад дробление яйца лошадиной аскариды (Ascaris megalocephala) (рис. 27). В этом случае происходит очень ранняя дифференциация ядер. В результате первого деления яйца, которое происходит в горизонтальной плоскости, анимальная его половина отделяется от вегетативной. В вегетативном бластомере хромосомы сохраняют вид двух длинных петель, какие были в яйце. В анимальном бластомере концы петель отбрасываются и впоследствии дегенерируют, а средние части хромосом распадаются на кусочки. При следующем делении вегетативного бластомера вновь в одном из продуктов деления сохраняются целые петлевидные хромосомы, а в другом — концы хромосом отбрасываются, а средние части распадаются. Так происходит в течение четырех последовательных делений, в результате которых получается зародыш, состоящий из 16 клеток, из которых только в одной сохраняются по две длинные хромосомы, а во всех других имеются многочислен-
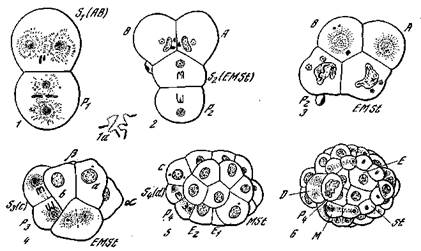
нее. 27. Дробление и диминуция хроматина в яйце Ascaris [Гексли и де Вер, W36]
1 — стадия двух бластомеров. В анимальном бластомере происходит диминуция хроматина; в вегетативном бластомере хромосомы сохраняют вид двух длинных Петель: la —диминуция хроматина (увеличено); 2 — Т-образная фаза; 3 — ромбоидная фаза; 4 — диминуция хроматина во втором соматобласте; 5, в — ^-клеточная и более поздняя стадии; Si (AB) — первый соматобласт (зачаток первичной эктодермы); s2 {EMSt) —второй соматобласт (энтомезостомодеальный зачаток); 4 (d) —зачатки вторичной и третичной эктодермы; Р4 — первичная половая клетка
300 Регуляция формообразования в индивидуальном развитии
ные мелкие хромосомы, получившиеся в результате распада средних частей длинных хромосом. В дальнейшем развитии клетка с длинными хромосомами переходит в состояние довольно длительного покоя. Это — первичная половая клетка. Обычные клетки энергично делятся и дают начало всему телу зародыша. Это — соматические клетки. Во время дробления у аскариды происходит, следовательно, дифференцировка ядер, которая определяет дальнейшую судьбу клеток в качестве половых (с полными хромосомами) или соматических (с диминуцией хроматина). Однако чем же определяется эта дифференциация ядер? Опыты центрифугирования яиц аскариды дают на это недвусмысленный ответ (рис. 28). При энергичном центрифугировании происходит более резкое разграничение субстанций плазмы, некоторое сплющивание яйца, в результате этого первая плоскость дробления проходит не горизонтально, а вертикально. Вегетативные и ани-мальные плазматические субстанции переходят одинаково в оба первых бластомера, и в обоих сохраняются неизменными обе длинные хромосомы. В дальнейшем из таких яиц развиваются уродливые двойники, содержащие пару половых зачатков. Эти опыты показывают вновь значение плазматических субстанций и их распределения для наступления известных дифференцировок. В данном случае в цитоплазме вегетативного полюса содержатся «коричневые зернышки», которые препятствуют распаду хромосом в тех бластомерах, в которые они попадают. Следовательно, хотя дифференциация ядер и определяет у аскариды образование половых и соматических клеток, она сама определяется цито-плазматическими субстанциями яйца.
Можно привести и, как будто, совершенно противоположные примеры. Мы вначале уже ознакомились с детерминированным развитием яиц асцидий, в которых дифференцировка частей зависит от перераспределения плазматических веществ, имеющихся уже в цитоплазме зрелого яйца. Последнее имеет сложную двустороннесимметричную структуру, которая окончательно устанавливается после оплодотворения.
Яйца кольчатых червей и моллюсков (рис. 29) также отличаются детерминированным дроблением и мозаичным развитием, зависящим от распределения различных цитоплазматических субстанций яйца. Структура яйца, устанавливающаяся еще в яичнике, однако, несимметричная. Дробление имеет характер спирального дробления, при котором веретена ядерных делений располагаются косо. Уже первые четыре бластомера несколько различаются по размерам и цитоплазматическим субстанциям. При третьем клеточном делении отделяется нижняя, вегетативная, половина каждого бластомера от верхней, анимальной. Так как веретена ядерных делений стоят не вертикально, а под некоторым углом, то и плоскость третьего дробления в каждом блас-томере лежит под углом к горизонтали. В результате этого деле-
Глава V. Взаимодействие между ядром и цитоплазмой 301
$ця. образуются квартет более крупных вегетативных клеток 0 лежащий над ним квартет более мелких анимальных клеток, g отличие от симметричных форм дробления анимальные блас-томеры лежат не прямо над соответствующими вегетативными бластомерами, а сдвинуты так, что располагаются против промежутков между ними. Если при взгляде со стороны анималь-ного полюса его клетки оказываются при третьем делении сдви-
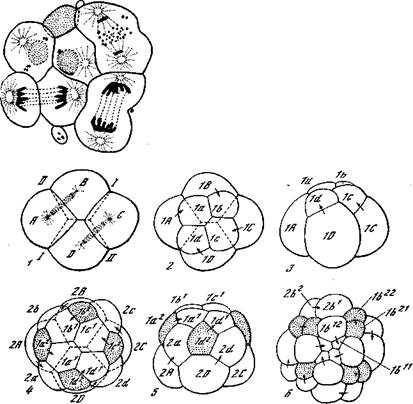
|
| Рис. 28. Дробление центрифугированного яйца Ascaris [Морган, 1927] Рис. 29. Спиральное дробление моллюска Trochus (Prosobranchia) [Кюн, 1955] 1 — 4-клеточная стадия; 2,8 — 8-клеточная; 4, 5 — 16-клеточная; 6 —32-клеточная. Точки — первичные трохобласты (4, 5) и их дочерние клетки (в). На последнем рисунке обозначены микромеры квадранта В |
нутыми по часовой стрелке от соответствующих вегетативных, то такое дробление называется дексиотропным, если же клетки анимального квартета сдвигаются против часовой стрелки по отношению к клеткам вегетативного квартета, то дробление называется леотропным. Как и при других типах дробления, положение плоскостей деления чередуется и каждая следующая плоскость располагается перпендикулярно к предыдущей. Поэтому и здесь после дексиотропного третьего деления следует лео-тРопное четвертое и т. д. В дальнейшем, по мере дифференциации продуктов деления, строгая последовательность делений
302 Регуляция формообразования в индивидуальном развитии
постепенно теряется и зародыш получает двусторонне-симметрич-ное строение, но все же спиральное дробление оставляет у большинства моллюсков ясный отпечаток на всей их организации, которая у них теряет свою симметрию вследствие преобладающего развития органов одной стороны тела. Это влечет за собой спиральное закручивание тела, что отражается и на внешней форме раковины, закрученной обычно вправо у форм
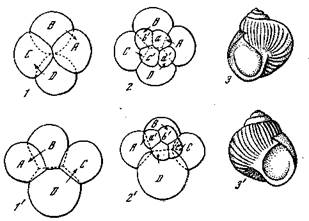
Рис. 30. Спиральное закручивание раковины Gastropoda {Морган, 1927]
1,2,3 — леотропное дробление и раковина, закрученная влево; 1', 2', 3' — дексио-тропное дробление и раковина, закрученная вправо
и дексиотропным дроблением и влево — у форм с леотропным дроЬлением (рис. 30). Обычно у гастропод наблюдается дексио-хропное дробление и раковины завернуты вправо. Гораздо реже встречаются виды с раковиной, закрученной влево. Интересно, что у видов с дексиотропным дроблением встречаются иногда и особи с раковинами, закрученными влево, которые передают это свойство по наследству как простые мутации, определяемые одним геном (Physa, Fraticicola lantzi). Как уже упомянуто, структура яйца моллюсков устанавливается еще в яичнике. Оказывается, у особей с раковинами, закрученными вправо, овоциты прикрепляются к стенке яичника своим вегетативным полюсом, а у леотропных особей — анимальным полюсом. Структура яйца определяется здесь, как и во многих других случаях, направлением поступления веществ из стенок яичника.
Таким образом, ясно, что дробление яйца и организация моллюска (дексио- или леотропное вращение) определяются распределением веществ цитоплазмы яйца, которое устанавливается под влиянием материнского организма. Однако вместе с тем эта
Глава V. Взаимодействие между ядром и цитоплазмой 303
организация наследуется как простой «менделирующий» признак. Форма наследования, правда, как будто, несколько измене-на? так как яйцо получает свою спиральную организацию только 0т матери. При скрещивании дексиотропных самок улиток с леотропными самцами (Physa, Limnaea) получаются дексиотроп-ные гибриды, откладывающие яйца с дексиотропным дроблением. В потомстве этих гибридов идет расщепление в отношении: три дексиотропных и одна леотропная особи. Расщепление запаздывает, следовательно, на одно поколение против обычного. Это вполне понятно, так как яйца первого поколения формировались под влиянием гомозиготной матери. Яйца второго поколения формировались под влиянием гетерозиготной матери, у которой были выражены лишь доминантные черты дексиотропной организации. Эти яйца получили также дексиотропную структуру, за которой скрываются, однако, различные генотипы {DD, Dd и dd). Рецессивная гомозигота и выявляется в третьем поколении — одна треть особей этого поколения, имея по матери раковины, завитые вправо, откладывает, однако, леотропные яйца, из которых развиваются моллюски с раковиной, закрученной влево. Следовательно, организация яиц определяется не фенотипом матери (имеющей дексиотропную организацию), а ее генотипом, т. е. структурой ее клеточных ядер (гомозиготных по вращению раковины влево — dd). Таким образом, один из лучших примеров, показывающих детерминирующее значение структуры яйца и его плазматических субстанций в формировании зародыша, доказывает вместе с тем с предельной ясностью, что эта структура яйца определяется строением ядер материнского организма, в котором это яйцо формируется. В данном случае, хотя дифференциация сомы и определяется плазматическими субстанциями яйца, строение последнего и его формообразующие вещества возникают под влиянием ядерных структур.
И во многих других случаях эмбрионального развития, в особенности на ранних стадиях, обнаруживается влияние материнского организма. Так, например, внешние свойства грены (форма и окраска яиц) тутового шелкопряда, а также скорость роста его гусениц обнаруживают явное влияние материнского организма. Однако и на этом объекте в последнее время В. Л. Астауровым доказано решающее значение ядерных структур. Ему удалось вывести у тутового шелкопряда не только вполне жизнеспособных гусениц, но и взрослых бабочек путем оплодотворения безъядерного яйца двумя сперматозоидами. Ядро яйца разрушалось действием повышенной температуры (или рентгеновыми лучами), а два ядра сперматозоидов сливались и Давали начало диплоидному ядру, которое обеспечивало совершенно нормальное развитие. Так как ядро каждого сперматозоида содержит у бабочек одну половую Z-хромосому, то все особи
304 Регуляция формообразования в индивидуальном развитии
получили по две Z-хромосомы и развились соответственно в самцов.
В этих опытах Б. Л. Астаурову удалась и гибридизация между домашним Bombyx mori и близким диким видом Вопь byx mandarina. При оплодотворении яйца Bombyx mandarina (ядро которого разрушено теплом) двумя сперматозоидами Bombyx mori развитие шло совершенно нормально — была получены взрослые плодовитые самцы со всеми признаками культурного вида. Обратное скрещивание удалось с рентгенизированными яйцами домашнего шелкопряда при оплодотворении сперматозоидами дикого вида. В этом случае получились гусеницы и коконы со всеми признаками дикого вида, однако взрослых бабочек получить не удалось. В обоих случаях все полученные гибриды обладали признаками того вида, от которого получено ядро. Плазма яйца не оказала никакого влияния на развитие. Так как различия между обоими видами шелкопрядов весьма значительны на всех стадиях развития, то в этом случае исключается возможность какой-либо ошибки.
Многочисленные эксперименты удаления ядра из яйцеклетки дали совершенно однозначные результаты — дробление яйца в некоторых случаях может совершаться и без ядра, однако на этом развитие и кончается. Формообразование без ядра, или, во всяком случае, без продуктов его деятельности (как у Acetabularia) — невозможно. Нормально дифференциров-ка частей зародыша определяется распределением и перераспределением субстанций яйца, а также образованием новых веществ в цитоплазме клеток зародыша. Всякая дифференцировка, орга-нотипичёская или гистологическая, начинается с изменений в составе цитоплазмы, т. е. с хемодифференциации.
Так как без ядра формообразование невозможно, то, очевидно (как это доказано для Acetabularia и для рассмотренных примеров развития яиц), эта дифференциация определяется ядром. Однако изменяется ли при этом и само ядро? А. Вейсман предполагал, что в основе явлений дифференцировки лежат именно качественно неравные деления ядер. Эта гипотеза была опровергнута, между прочим, следующим красивым экспериментом (рис. 31). Яйцо тритона перетягивалось тонким волосом приблизительно по плоскости симметрии (чтобы разделить серый полумесяц) так, что оно приняло форму гантели, в одной половине которой помещалось ядро. В этом случае дробилась только половина, содержащая ядро. После образования в этой половине 8 или 16 клеток петля волоса несколько ослаблялась и ядро ближайшей клетки проникало в нераздробившуюся половину яйца. После этого петля затягивалась до конца и отделяла раздробившуюся половину с 7 или 15 ядрами от второй, нераздробившейся половины. После этого происходило дробление второй половины яйца и, конечно, продолжалось развитие и первой половины.
Глава V. Взаимодействие между ядром и цитоплазмой 305
g результате в одной оболочке, получались две целых, совершенно нормальных зародыша и вылуплялись одинаковые нормаль-яые личинки. Из этих личинок одна получила 15 ядер 16-кле-точной стадии, а другая только одно из ее ядер. Следовательно, на стадии 16 бластомеров все ядра еще совершенно одинаковы, каждое из этих ядер может обеспечить развитие целого, вполне
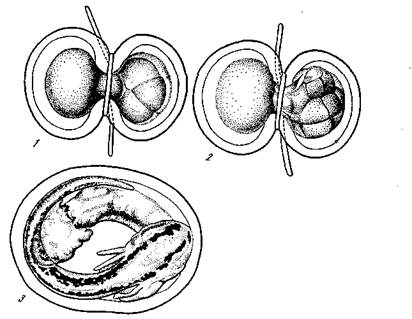 Рис. 31. Перешнуровка яйца Triton taeniatus на стадии 16 бластомеров [Гексли и де Бер, 1936]
Рис. 31. Перешнуровка яйца Triton taeniatus на стадии 16 бластомеров [Гексли и де Бер, 1936]
1 — оплодотворенное яйцо перетянуто лигатурой, благодаря чему ядро оказалось в правой половине, левая половина не дробится;
2 — на стадии 16 бластомеров одно из 16 ядер проникает в левую,
не раздробившуюся половину, и туго затянутая лигатура полностью разделяет обе половины яйца; 3 — каждая половина яйца развилась в нормального зародыша
нормального организма. Это послужило ясным опровержением гипотезы неравноценных делений ядра. Ядра испытывают в процессе митотического деления всегда равнонаследственное распределение материала.
И однако мы знаем, что ядра могут испытывать различную Дифференцировку. Мы уже привели яйцо аскариды как пример зависимой от цитоплазмы диминуции хроматина в соматических клетках. Прямые наблюдения показывают, что в некоторых тканях насекомых происходит вполне закономерное увеличение числа наборов хромосом (при разделении хромосом без Деления клетки — эндомитоз) — полиплоидия. Иногда происходит скрытое увеличение числа наборов хромосом, не отделяющихся друг от друга — политения. Таковы, например, гигантские хромосомы в ядрах клеток слюнных желез насекомых. Во
306 Регуляция формообразования в индивидуальном развитии
всех этих случаях изменения в ядрах наступают в результате дифференцировки клеточной плазмы. Однако несомненно, что такие измененные ядра оказывают самое непосредственное влияние на клеточный метаболизм, а следовательно, и на дальнейшие процессы дифференцировки. В процессах клеточной дифференцировки большое значение имеют, по-видимому, и хромосомные перестройки и определенные изменения в механизме митотического деления (многополюсные митозы). Во всяком случае, это показано для клеток железистых и особенно кроветворных органов млекопитающих.
Если ядра в яйце тритона все еще равнозначны на стадиях 8 или 16 бластомеров, то это, очевидно, связано с тем, что на стадиях дробления нет еще никакого формообразования, никакой дифференцировки. Формообразование начинается у амфибий с гаструляции. Если ядра дифференцируются по-разному в разных частях зародыша, то этого можно ожидать именно на стадиях гаструляции. Это предположение проверялось методом пересадки ядер. У предварительно активированного яйца лягушки удаляли ядро и после этого микропипеткой вносили в него ядро, извлеченное из клетки определенной области бластулы или гаструлы. Ядра клеток анимальной половины бластулы или ранней гаструлы дают при пересадке в энуклеированное яйцо нередко вполне нормальное развитие целой личинки. Следовательно, на стадии ранней гаструлы ядра во всяком случае еще равноценны. На стадиях поздней гаструлы пересадка ядер встречала более значительные технические трудности, после преодоления которых (разделение клеток с помощью трипсина и версена) удалось получить следующие результаты. Ядра, выделенные из хордомезодермы поздней гаструлы и внесенные в энуклеированное яйцо, вызывали нормальное дробление, а иногда и гаструляцию. Но на стадии нейрулы развитие прекращалось. В таких зародышах хорда и сомиты были нормальны, но нервная система недоразвита. При такой же имплантации в яйца лягушки ядер, извлеченных из энтодермы дна первичной кишки, наблюдалась остановка развития на разных стадиях бластулы, гаструлы или нейрулы. Развивались аномальные зародыши с нормальной хордой и кишечником, крупными сомитами аномальной формы и недоразвитой эктодермой.
Эти опыты, как будто, доказывают, что на стадии поздней гаструлы, т. е. именно тогда, когда у амфибий завершается детерминация основных частей зародыша, дифференцируются и ядра. Ядра, извлеченные из клеток одного первооргана, в результате предшествовавшего влияния дифференцированной уже цитоплазмы, по-видимому, теряют способность обеспечить нормальное развитие других частей зародыша, хотя они и попадают в нормальную цитоплазму со всеми необходимыми для этого субстанциями. Однако возможно, что эти результаты объясняют-
Глава V. Взаимодействие между ядром и цитоплазмой 307
ся повреждением ядер при их изоляции на более поздних стадиях развития.
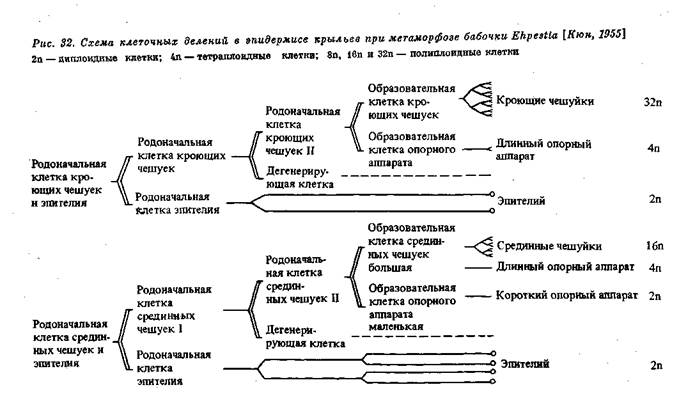
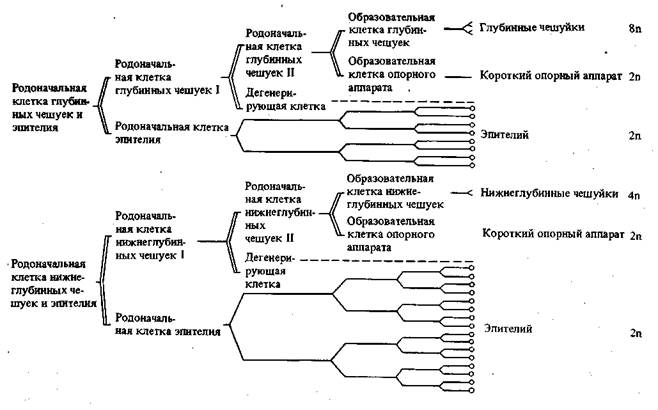
В некоторых случаях, как уже упомянуто, влияние плазматической дифференцировки сказывается и в видимом изменении структуры ядра, а в результате и в дальнейшей судьбе соответствующих клеток. При дифференцировке эпидермиса в куколке бабочек родоначальные клетки — образователи чешуек — испытывают совершенно определенное число делений, дающих начало, с одной стороны, обычным эпителиальным клеткам и, с другой стороны, клеткам — образователям чешуек (рис. 32). В первых происходят обычные митотические деления, а во вторых — такое же число эндомитотических делений ядер, приводящих к образованию полиплоидных клеток разной плоидности. Здесь разная плоидность связана с разной величиной и разным положением развивающихся чешуек (рис. 33).
Опыты с пересадками ядер должны дополнить наши знания о дифференцировке ядер в процессах формообразования. Однако все известные до сих пор факты дают вполне определенные указания на то, что формообразование вообще возможно лишь на основе взаимодействия между ядром и плазмой. Клеточная и органная дифференцировка начинается всегда с изменений в цитоплазме (хемодифференциация). Если наблюдается дифференцировка ядер, то это является следствием наступившей уже дифференцировки клеточных тел. С другой стороны, специфические особенности плазматических дифференцировок и конкретная реализация всего формообразования неизменно определяются наследственными свойствами клеточных ядер или, точнее, их взаимодействием с цитоплазмой, так как без этого взаимодействия никакие жизненные функции невозможны. Эти выводы подкрепляются также огромным арсеналом фактов, добытых при изучении развития организмов с измененными соотношениями между клеточными ядрами и цитоплазмой.
При этом во всех случаях совершенно ясно проявляется специфика влияния ядерных субстанций. Остается, однако, вопрос, как это влияние осуществляется? Мы уже неоднократно подчеркивали, что это происходит через посредство рибонуклеиновых кислот (РНК), синтез которых определяется кодом ядерной ДНК и которые, в свою очередь, определяют синтез специфических белков. Механизм реализации этих процессов в известной степени уже выяснен. Связь между синтезом белков и кодом ДНК (через посредство РНК) вполне определяет структуру белков и последовательность входящих в них аминокислот. Каждой из Имеющихся 20 аминокислот соответствует определенное сочетание трех (из четырех возможных) нуклеотидов цепи молекулы ДНК. В наследственном коде ДНК каждый последовательный Триплет определяет синтез известной аминокислоты, и весь непрерывный ряд нуклеотидов определяет, таким образом, соответ»
|
|
310 Регуляция формообразования в индивидуальном развитии
ствующий ряд аминокислот. Наследственные структуры ядра (ДНК) реализуются, следовательно, в индивидуальном развитии через посредство синтеза активных субстанций протоплазмы —. рибонуклеиновых кислот (РНК) и белков. Поступление этих субстанций в цитоплазму можно иногда непосредственно наблюдать. Давно уже известно, что при образовании яйца в яичнике (в фазе роста) хромосомы принимают вид «ламповых щеток».
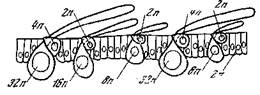
Рис. 33. Схема полиплоидии ядер в эпидермисе крыльев бабочки [Кюн, 1955]
Кроющие чешуйки (32 п); срединные чешуйки (16 п); глубинные чешуйки (8 п); опорный аппарат чешуек (2 п и 4 п) и клетки эпидермиса (2 п)
Оказывается, во всех поперечных выростах хромосом содержится рибонуклеиновая кислота, синтезируемая в определенных точках хромосом (соответственно положению некоторых активированных генов). Эта РНК поступает в цитоплазму, таким образом яйцо нагружается запасом активных веществ, специфика которых определяется генотипом неоплодотворенного яйца. Это объясняет материнское наследование некоторых особенностей ранних стадий развития (например, ассиметрию дробления у моллюсков).
Еще интереснее наблюдения над поведением гигантских хромосом слюнных желез у личинок двукрылых (в частности, у классического объекта генетических исследований — плодовой мушки дрозофилы). Во время развития в хромосомах выявляются особо активные сильно вздутые участки — «пуффы», на которых опять-таки происходит интенсивный синтез РНК, поступающий затем в цитоплазму. Особенно интересно, что на разных стадиях развития функционируют различные пуффы. При переходе от одной стадии к другой одни пуффы исчезают, а другие возникают. Порядок возникновения пуффов специфичен для данного вида, но при мутациях может измениться. Следовательно, он определяется строением наследственного кода. Появление пуффов означает активацию некоторых генов, которые выделяют свою продукцию в цитоплазму и определяют тем самым ход ее диф-ференцировки. В этом выражается влияние наследственной структуры ядра на процессы развития. С другой стороны, однако, последовательная активация генов и возникновение новых пуффов происходят под влиянием продуктов дифференцировки, т. е. предшествующего этапа деятельности ядра. Такое влияние ока-
Глава VI. Ядро и плазма 6 нарушенных системах 311
зывают и гормоны. Экспериментально доказано (в эксплантатах), что под действием гормона метаморфоза в хромосомах слюнных ^елез личинок плодовой мушки возникают те же самые пуффы, которые характерны для периода метаморфоза в нормальном развитии.
Во всех этих случаях совершенно ясно обнаруживается взаимодействие между ядром и цитоплазмой в виде замкнутого цикла зависимостей. Активированные гены, т. е. участки хромосом ядра, определяют образование продуктов дифференци-ровки цитоплазмы (реализация наследственной информации), а продукты дифференцировки цитоплазмы, в свою очередь, определяют активацию известных участков хромосом, т. е. дальнейших генов (обратная связь). Все это показывает, что дифференцировка ядер во время развития состоит в последовательной и, очевидно, обратимой активации некоторых генов.
Глава VI
ВЗАИМОДЕЙСТВИЕ
МЕЖДУ ЯДРОМ И ЦИТОПЛАЗМОЙ
В НАРУШЕННЫХ СИСТЕМАХ
(ГИБРИДЫ И МУТАЦИИ)
Если формообразование определяется взаимодействием частей с различно дифференцированной цитоплазмой, а эта дифференцировка, в свою очередь, является выражением взаимодействия между ядром и цитоплазмой, то естественно ожидать что при совмещении ядра и цитоплазмы разных видов или при изменении строения ядра и его хромосом нормальное формообразование окажется нарушенным. При межвидовой гибридизации всегда происходит совмещение цитоплазмы яйца одного вида с ядром, имеющим смешанное происхождение от обоих видов. Результат зависит от степени межвидовых различий. Более отдаленные формы вообще не дают гибридов, так как их половые клетки несовместимы. Так как со сперматозоидом вносится лишь ничтожная масса цитоплазмы, то мы можем с достаточным основанием говорить о несовместимости ядра сперматозоида с цитоплазмой яйца другого вида. Это подтверждается многочисленными наблюдениями, когда в результате экспериментальной гибридизации яйцо активируется и приступает к нормальному дроблению, но при участии одного только материнского ядра. Ядро, внесенное сперматозоидом, остается неактивным и постепенно дегенерирует. В этих случаях яйцо развивается по сути партеногенетически
312 Регуляция формообразования в индивидуальном развитии
(гиногенез) и может дать начало нормальному гаплоидному организму. Гибель ядра сперматозоида указывает на полную невозможность взаимодействия между ядром и цитоплазмой, являющегося основой жизненных процессов любой клетки.
В других случаях, а именно тогда, когда видовые различия не столь велики, объединение обоих ядер происходит нормально. Яйцо дробится, развитие зародыша начинается, как будто, нормально, но рано или поздно останавливается, и зародыш гибнет. Ясно, что взаимодействие между хромосомами отцовского ядра и материнской цитоплазмой все же нарушено.
В последнее время подробно изучены различные комбинации межвидовых гибридов у амфибий. Обычно такая гибридизация имеет летальный эффект даже в случае развития гаплоидного организма. Обычно развитие гибридов бесхвостых амфибий останавливается на стадии поздней бластулы или ранней гаструлы, т. е. как раз тогда, когда начинаются процессы детерминации частей зародыша, связанные с изменением клеточного метаболизма. Формообразование возможно лишь на основе нормального взаимодействия между ядром и цитоплазмой одного и того же вида. У гибридов это взаимодействие оказывается нарушенным.
У хвостатых амфибий гибриды, как правило, живут дольше, но все же обычно до метаморфоза не доходят. Даже такие близкие виды, как Triturus cristatus и Triturus marmoratus, которые естественно спариваются в местах совместного обитания (Западная Франция), дают гибридов, которые в большинстве гибнут на личиночных стадиях развития и не доходят до метаморфоза. Лишь очень немногие гибриды проходят благополучно метаморфоз, а еще реже они достигают половозрелости. Естественные гибриды этих видов были описаны как Triturus blasii. Численное соотношение полов у них несколько нарушено в пользу самок. Плодовитость последних при скрещивании с исходными видами сильно снижена, а самцы совсем стерильны. Все это говорит о глубоких нарушениях развития даже при гибридизации между очень близкими видами. В некоторых случаях гибридизация возможна и между ясно обособленными видами, однако такие гибриды обычно бесплодны (классический пример — мулы). Бесплодие гибридов объясняется глубокими нарушениями в процессах созревания половых клеток. Эти нарушения являются особенно чувствительным реагентом. Даже при скрещивании подвидов или различных рас плодовитость полученных гибридов, несмотря на их зачастую мощное развитие (гетерозис), обычно оказывается сниженной. Это всегда указывает на различное строение основных компонентов ядра — хромосом, которые образуют в каждом виде и расе определенную точно сбалансированную систему. При совмещении хромосом двух разных видов или даже рас в одном ядре этот нормальный генный баланс оказывается нарушенным. Это сказывается, прежде всего, на плодо-
Глава VI. Ядро и плазма в нарушенных системах 313
вцтости, а затем, при больших различиях, и на жизнеспособности развивающихся гибридов.
Значение ядерных структур (в системе ядро — цитоплазма) для развития видно также при сопоставлении организмов с измененным числом наборов хромосом (от гаплоидов до полиплоидов), с измененным числом хромосом в отдельном наборе (ге-тероплоидия), с измененной структурой хромосом (перестройки и нехватки) и с измененными элементарными их единицами — генами (генные мутации). Эти случаи особенно интересны для разбираемых нами вопросов формообразования, так как при этом происходит не сопоставление ядра и цитоплазмы разных видов или рас, т. е. чуждых образований, а сопоставление измененного ядра с цитоплазмой того же вида. В этом случае не может быть речи о биохимической несовместимости веществ цитоплазмы и ядра, а только о нарушении нормальных соотношений.
Простые нарушения количественных соотношений между нормальным ядром и цитоплазмой, как и нужно было ожидать, не ведут к каким-либо значительным изменениям. Раньше этим соотношениям приписывалось даже известное детерминирующее значение. Однако экспериментальное нарушение чисто количественных соотношений ведет к быстрому их восстановлению. При искусственном партеногенезе яйца морского ежа в нормальной массе его протоплазмы заключается вместо двух ядер только одно материнское ядро и, следовательно, половина нормального числа хромосом. Такое гаплоидное яйцо дробится совершенно нормально. При каждом делении масса цитоплазмы уменьшается примерно вдвое. Гаплоидное яйцо испытывает, однако, еще одно лишнее деление, в результате этого масса цитоплазмы становится в каждой клетке вдвое меньше, чем в норме, т. е. в точности соответствует своим уменьшенным ядрам. Число клеток соответственно оказывается вдвое большим, хотя зародыш имеет нормальную величину. При экспериментальном получении тетраплоидных зародышей (через подавление первого деления цитоплазматического тела яйца) их ядра содержат вдвое большее число хромосом и имеют, соответственно, вдвое больший объем при той же массе протоплазмы, как и в нормальных диплоидных клетках. В этом случае число клеточных делений сокращается. Получаются опять вполне нормальные зародыши с вдвое большими клетками, но с вдвое меньшим числом клеток. С другой стороны, можно искусственно Уменьшить массу цитоплазмы — просто разделить яйцо на части. Часть, содержащую ядро, можно оплодотворить. В этом случае Дробление заканчивается раньше нормы, но вновь именно тогда, Когда установится нормальное соотношение между объемами яДра и цитоплазмы. Так как ядро имеет в этом случае объем нормального диплоидного ядра, то и клетки получаются нормально-
314 Регуляция формообразования в индивидуальном развитии
го размера. Однако число клеток зародыша оказывается уменьшенным соответственно уменьшенной массе яйца. Соответственно оказывается уменьшенной и величина зародыша. Таким образом, в результате дробления в клетках зародыша всегда устанавливается нормальное объемное соотношение между ядром (точнее, числом наборов его хромосом) и цитоплазмой.
Мы видим, что чисто количественные соотношения между ядром и цитоплазмой хорошо регулируются. Нормальные соотношения легко восстанавливаются, а формообразование не нарушается. Однако формообразование, конечно, нарушается при качественных изменениях в составе цитоплазмы яйца (например, при изъятии некоторых активных ее веществ) или в составе хромосом его ядра. В некоторых случаях, однако, и чисто, казалось бы, количественные изменения в ядре вносят все же некоторые изменения, если не прямо в формообразование, то в процессы обмена веществ, отражающиеся на жизнеспособности, устойчивости развивающихся особей или на их плодовитости.
Так, например, гаплоиды, получаемые при искусственном партеногенезе, обычно менее жизнеспособны, чем нормальные диплоидные особи. В особенности это касается позвоночных животных. В этом случае, однако, не следует думать, что гаплоидный организм отличается от диплоидного всегда только по количеству хромосом. Гомологичные хромосомы никогда не бывают вполне идентичными, в особенности у высших животных с двуполым размножением. В настоящее время этому придается большое значение — известная степень генных различий между гомологичными хромосомами (т. е. известный уровень гетерози-готности) оказывается существенным условием для развития вполне жизнеспособного и устойчивого организма с высокой ре-гуляторной способностью (т. е. с совершенными гомеостатичес-кими механизмами развития). Организмы с совершенно одинаковыми наборами хромосом оказываются менее жизнеспособными и малоустойчивыми. Неудивительно, что и особи с одним набором хромосом мало жизнеспособны. С другой стороны, особи с увеличенным числом наборов хромосом (полиплоиды) не только не обнаруживают особого нарушения формообразования, но у растений нередко выделяются высокой жизнеспособностью и устойчивостью. (Это не значит, что формообразование вовсе не изменяется. Мы знаем, что даже простое изменение массы зачатка ведет к изменению скорости дифференцировки и ее полноты. И полиплоиды отличаются иногда своими размерами и морфологическими признаками. Кроме того, они обладают сниженной плодовитостью вследствие нарушений в делениях созревания половых клеток.)
В то же время развитие яиц, обладающих, кроме нормального диплоидного набора, еще одной или двумя добавочными хромосрмами, оказывается у высших животных, как правило,
Глава VI. Йдро и плазма в Нарушенных системах 315
нарушенным. Формообразование идет ненормально, жизнеспособность зародыша сильно снижена. При неправильном распределении хромосом во время дробления яиц с лишними хромосомами развитие не идет дальше бластулы. Нехватка отдельных хромосом в большинстве случаев также летальна. Для нормального развития яйца требуется, следовательно, наличие нормального сбалансированного набора хромосом. Только во взаимодействии такого эуплоидного ядра с соответствующей ему цитоплазмой возможны нормальный клеточный метаболизм и нормальное формообразование, приводящие к развитию вполне жизнеспособного и устойчивого организма.
Как велико значение ядерных структур в индивидуальном развитии, видно в особенности ясно при изменениях в строении отдельных хромосом. Мы знаем различные типы таких изменений. Самым обычным является обмен между соответствующими частями гомологичных хромосом (перекрест). Нередко наблюдаются перестановки и повороты частей в пределах одной хромосомы. Такие изменения не влияют на формообразование (за немногими исключениями, когда сказываются взаимовлияния между соседними генами—«эффект положения»). Бывает перенос части одной хромосомы на другую. Такие преобразования также не вносят заметных нарушений в формообразование. Иногда часть хромосомы переносится на гомологичную хромосому (без обмена). Тогда одна хромосома имеет удвоенную часть (дупликация), а у гомологичной хромосомы этой части нет вовсе (нехватка). В потомстве таких особей могут получиться при оплодотворении такими же сперматозоидами яйца с удвоением или с нехватками в обеих гомологичных хромосомах. Дупликации обычно не влияют на развитие существенным образом, особенно в гетерозиготном состоянии. Однако все же их жизнеспособность бывает сниженной. Некоторые дупликации оказываются даже летальными. В этом сказывается царушение генного баланса. Однако нехватки в гомозиготном состоянии уже, как правило, легальны. Даже гетерозиготные нехватки связаны со снижением жизнеспособности и часто детальны. Только очень небольшие нехватки проявляются у дрозофилы как жизнеспособные в гетерозиготном состоянии мутации. В этом отношении они сходны с точечными, или генными мутациями. Предполагайся, что некоторые летальные генные мутации в действительности представляют собой такие малые нехватки.
Как мы видим, нарушение грубого строения хромосом либо Не влияет на формообразование (если нормальный баланс не Нарушен), либо резко сказывается на жизнеспособности развивающегося организма (если нарушены количественные соотношения внутри ядра, т. е. генный баланс, и в особенности при Нехватках). Для разбираемых нами вопросов гораздо интереснее качественные изменения хромосом, захватывающие отдель-
316 Регуляция формообразования в индивидуальном развитии
ные гены (точечные мутации). Не нарушая баланса и не имея обычно летального влияния, они нередко заметно изменяют клеточный метаболизм и в результате этого вносят существенные изменения в процессы формообразования.
Генные мутации в настоящее время рассматриваются как изменения в химическом строении молекулы дезоксирибонуклеино-вой кислоты. Каждая мутация есть результат некоторой перестройки этой молекулы, состоящей в изменении расположения пар оснований (аденин с тимином и цитозин с гуанином), хотя бы в одном только звене. В таком случае следует ожидать, что мутации вызываются химическими агентами и выражаются, прежде всего, в изменении клеточного метаболизма и в изменении его продуктов.
В пользу предположения о химической природе мутаций говорят очень многие факты:
1. Возможность экспериментального получения мутаций под непосредственным влиянием разнообразных химических агентов.
2. Прямая пропорциональность мутагенного действия ионизирующей радиации применяемой дозе, т. е. числу ионизации. Ионизация сопровождается образованием перекиси водорода, которая является мутагенным фактором.
3. Возрастание частоты спонтанных мутаций с повышением температуры соответственно (в известных пределах) закону Вант-Гоффа для химических реакций.
4. Увеличение числа мутаций по мере пребывания спермы в семяприемниках самки (у дрозофилы) и при хранении семян растений. Это указывает на возможное влияние нормальных метаболитов.
5. Значительное увеличение числа мутаций под влиянием
теплового шока (несомненное изменение метаболизма).
6. Мутагенное действие ультрафиолетовых лучей, дающих максимальное число мутаций на волне 2600 А. На этой же волне ультрафиолетовые лучи максимально поглощаются дезоксирибо-нуклеиновой кислотой.
7. Трансформирующее влияние измененной ДНК (от мутировавшего штамма) на нормальный генотип, установленное у бактерий. По-видимому, это связано с прямым вхождением молекул ДНК или их обломков в состав генотипа.
Если молекула ДНК прямо или косвенно контролирует кле4 точный метаболизм, то мутация должна прежде всего выражаться в химическом изменении его продуктов. У наиболее простых организмов — бактерий и низших грибов это как раз и выявляется с предельной ясностью. У растений хорошо изучено наследование окрасок цветов и плодов. Мутации связаны здесь с изменением химического строения растворенных в клеточном соке пигментов — антоцианов и антоксантинов, которые образуются из цианидина в различных условиях окисления или восста-
Глава VI. Ядро и плазма в нарушенных системах 317
яовления. Различные каротиноиды, образующиеся в пластидах и определяющие окраску плодов, листьев и других органов растений (от желтой до красной), также легко перестраиваются, и эти химические перестройки связаны с простыми мутациями. Наследование количественных и качественных различий углеводов, белков, жиров и витаминов в семенах растений обнаруживает закономерности, характерные для простых мутаций. Мутации обнаруживают и антигенные различия белков. Внутривидовые различия в антигенах на поверхности эритроцитов крови человека, выявляющиеся в известных группах крови (А, В, АВ и О), обусловлены серией аллеломорфных генов из трех членов. Высокая яйценоскость кур оказалась связанной с определенным комплексом антигенов.
Пигментообразование является и у животных чисто химической реакцией и, вместе с тем, находится в прямой зависимости от определенных генов. Однако окраска животных всегда связана и с более сложными процессами, определяющими распределение пигментов. Последнее также контролируется генами (как мы уже видели на очень простом еще примере аксолотля). У человека известно довольно много мутаций, связанных с изменением строения гемоглобина. Это типичные примеры химических мутаций, проявляющихся в виде наследственных заболеваний крови. Интересны мутации серповидноклеточной анемии негров и талассемии населения средиземноморских стран. Эти мутации в гомозиготном состоянии детальны, а в гетерозиготном состоянии вызывают анемию. Эти мутации, несмотря на их вредность, широко распространены в малярийных местностях. Оказывается, они связаны с устойчивостью против малярии, что и способствует их сохранению под влиянием естественного отбора. Серповидноклеточность определяется изменением одного только гена и выражается в образовании измененного гемоглобина, в котором одна из многих (около 300) аминокислот — глутаминовая кислота — замещена другой — валином.
Если мутации являются результатом перестройки молекулы ДНК, то это связывается и с изменениями в зависимых от нее структурах рибонуклеиновых кислот, а следовательно, и в составе активных белков — ферментов (которые синтезируются при посредстве РНК). Таким образом, вполне понятно, что каждая мутация выражается в изменении клеточного метаболизма и его продуктов. Следовательно, изменяются именно те клеточные субстанции, которые, как мы видели, определяют формообразование и дифференцировку частей зародыша и его клеток. Изменение клеточного метаболизма связано, конечно, и с изменением форм реагирования на различные внутренние и внешние факторы. Соответственно меняются и движения клеточных масс и миграции отдельных клеток. При мутации мохно-Ногости у кур скелетогенная мезенхима у заднего края лапки
с
318 Регуляция формообразования в индивидуальном развитии
вместо концентрации в зачат








