 2015-05-06
2015-05-06 538
538азотосодержащих соединений [41, 49, 223]
Эти последовательности изображены на рис.12. Реакции азотсодержащих соединений характерны для последовательностей только этого типа. Их повторяющиеся интервалы состоят из пяти реакций. Эти реакции дезаминирования или переаминирования a-аминокислот в a-кетокислоты, декарбоксилирования a-аминокислот в амины, окисления аминов в альдегиды, конденсации альдегидов с глицином в a-амино-b-оксикислоты и, наконец, дезаминирования последних в a-кетокислоты. Все превращения протекают с участием пиридоксальзависимых ферментов. Реакции дезаминирования аминокислот и аминов могут также осуществляться ферментами, зависимыми от окислительно-восстановительных коферментов. Реакции дезаминирования a-амино-b-оксикислот и декарбоксилирования a-аминокислот [41 с. 391] необратимы. В последовательностях этого типа отсутствуют вырожденные интервалы.
Нулевой инткрвал последовательности М состоит из реакций, связыающих глиоксилат и пируват. Реакция конденсации глицина с форальдегидом в серин, которая является промежуточной в этом интерале, имеет ту особенность, что формальдегид участвует в ней в форме соединения с фолиевой кислотой [260].
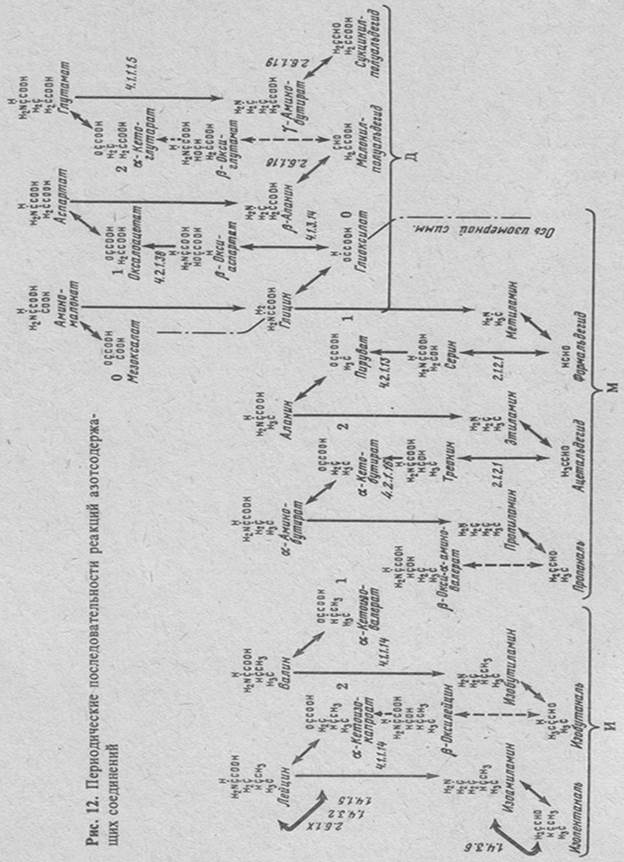
Первый интервал последовательности М состоит из реакций, связывющих пируват и a-кетобутират. Реакции этого интервала в биохимии рассматриваются в связи с метаболизмом аланина и треонина [113, 150, 190, 260].
Второй интервал последовательности М связан с метаболизмом непротеиногенной a-аминомасляной кислоты [41, 170]. К этому же интервалу должен быть отнесен выделенный из растительных объектов b-оксинорвалин [41 с. 308] метаболизм которого в настоящее время не изучен.
Нулевой интервал последовательности Д состоит из реакций, связыающих мезоксалат и оксалоацетат. В качестве альдегида в этом интервале выступает глиоксилат, который в нулевом периоде последовательности М играл роль исходной кетокислоты. Начальные реакции этого интервала обнаружены у шелковичного червя и в печени крысы [78, 235].
Первый интервал последовательности Д начинается реакциями аминирования оксалоацетата в аспартат, декарбоксилирования аспартата в b-аланин и дезаминирования b-аланина в малонилполуальдегид. Две последующие реакции этого интервала - образование b-оксиглутамата при конденсации малонилполуальдегида с глицином и дезаминирование b-оксиглутамата в a-кетоглутарат - в настоящее время неизвестны и могут лишь предполагаться, исходя из факта обнаружения производных b-оксиглутамата в некоторых биологических объектах [41б с. 351].
Аналогично неполными являются второй и третий интервалы данной последовательности. Реакции этих интервалов связаны с обменом соответственно глутаминовой и a-аминоадипиновой кислот [205, 256].
Путь расщепления глутамата во втором интервале, включающий его декарбоксилирование в g-аминомасляную кислоту и дезаминирование последней в сукцинилполуальдегид, характерен для тканей мозга и рассматривается как шунтовой по отношению к пути расщепления через a-кетоглутарат в цикле Кребса [75].
Переходная последовательность на участке связи последовательностей М и Д азотсодержащих соединений состоит всего лишь из двух реакций: декарбоксилирования аминомалоната в глицин [235] и декарбоксилирования глицина в метиламин [41 с. 391].
Изомерная симметрия связи между последовательностями М и Д проявляется в двух обобщенных рядах: в ряду a- и w-аминокислот и в ряду a-кето и w-альдегидокислот. Левые и правые участки ряда a- и w-аминокислот объединяются глицином. Соответствующие участки ряда a-кето и w-альдегидокислот - глиоксилатом.
Так же как и в высших интервалах последовательностей М и Д, реакции превращений b-окси-a-аминокислот в последовательностях И и И' в настоящее время неизвестны. Поэтому в первом интервале последовательности И на рис.12 приведен лишь встречающийся в природе b-оксилейцин [186], но не реакции его превращений. Второй интервал этой последовательности представлен частично: в нем отсутствует соответствующая a-амино-b-оксикислота и ее реакции.
Связь между последовательностями И и И" и последовательностью М на рис.12 не показана. Она может осуществляться через a-кетокислоты посредством тех же реакций, что и в последовательностях, описанных в разделе 4.1. Соответственно симметрия взаимосвязи этих последовательностей тоже сохраняется.








