 2015-04-20
2015-04-20 3330
3330Сенсорные (афферентные) нейроны (СН) - sensory (afferent) neurons - передают информацию о внешней или внутренней среде в обрабатывающие центры. Первичные СН находятся вне ЦНС, по строению относятся к униполярным, СН более высокого уровня находятся в ЦНС и относятся к мультиполярным.
Моторные (эфферентные) нейроны (МН) - motor (efferent) neurons - передают управляющие влияния от обрабатывающих центров к эффекторам (мышцам, железам). Тела МН находятся внутри ЦНС. МН высшего порядка принадлежат мозгу, отростки МН низшего порядка покидают мозг и относятся к периферической нервной системе. По строению МН являются мультиполярными. Вставочные (ассоциативные) нейроны (ВН) - inter (association) neurons - интегрируют информацию, поступающую в ЦНС осуществляют взаимодействие между сенсорной и моторной частями НС. Тела ВН находятся внутри ЦНС. По строению ВН являются анаксонными или мультиполярными.
Аня: По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на комиссуральные и проекционные (головной мозг).
Изменения эффективности синаптических контактов в нейронных сетях при длительной стимуляции: синаптическая потенциация и синаптическая депрессия, их роль в приспособительном поведении.
Аня: Долговременная синаптическая пластичность
Впервые идея о связи между получением опыта и изменением синаптической силы была высказана на рубеже XIX и ХХ вв. нобелевским лауреатом Сантьяго Рамон-и-Кахалем. Экспериментальное изучение долговременной синаптической пластичности базируется на постулате Хэбба, сформулированном в 1949 году: «Если аксон клетки А расположен достаточно близко к клетке Б, чтобы возбуждать её, и постоянно участвует в её активации, то в одной или обеих клетках происходят такие метаболические изменения или процессы роста, что эффективность А как одной из клеток, активирующих Б, повышается»[5]. В современной формулировке постулат Хэбба понимается так, что изменение эффективности передачи сигнала в синапсе управляется корреляцией силы, необходимой для активации пре- и постсинаптического нейрона[6].
Первые экспериментальные результаты, подтверждающие постулат Хэбба, были получены в начале 1970-х годов[7]: многократная активация возбуждающих синапсов гиппокампа кролика вызвала увеличение силы синапса, продолжавшееся несколько часов или даже дней. Это явление получило название «долговременная потенциация» (long-term potentiation, LTP). Позже были открыты другие явления, связанные с синаптической пластичностью, — долговременное ослабление (long-term depression, LTD), гомеостатическая пластичность, метапластичность. Гомеостатическая пластичность, например, представляет собой изменение силы всех синапсов конкретной клетки в ответ на длительные изменения активности, в частности, увеличение силы синапсов в ответ на уменьшение активности сигналов. Этот вид пластичности связан с гораздо большими временами, чем LTP и LTD, и может быть важен при развитии нейрональных путей. Термин «метапластичность» относится к эффектам, связанным с изменением возможности синапса проявлять пластичность[1]. В отличие от кратковременной пластичности, долговременная пластичность связана с экспрессией генов и синтезом новых белков. Наиболее изученными формами долговременной пластичности по-прежнему остаются LTP и LTD в области гиппокампа СА1, которые управляются N-метил-D-аспартатными (NMDA) рецепторами. Лишь в последние годы были получены доказательства в пользу существования долговременной потенциации ГАМКА-опосредованной синаптической передачи (LTPGABA), происходящей по гетеросинаптическому механизму, но и этот процесс требует активации NMDA-рецепторов в глутаматных синапсах[8].
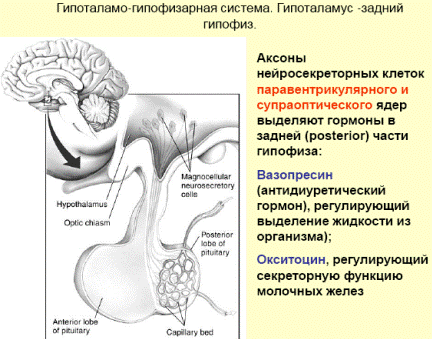
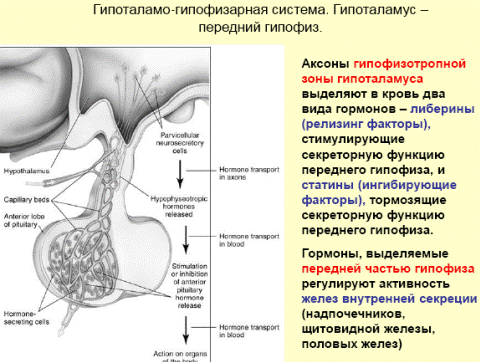
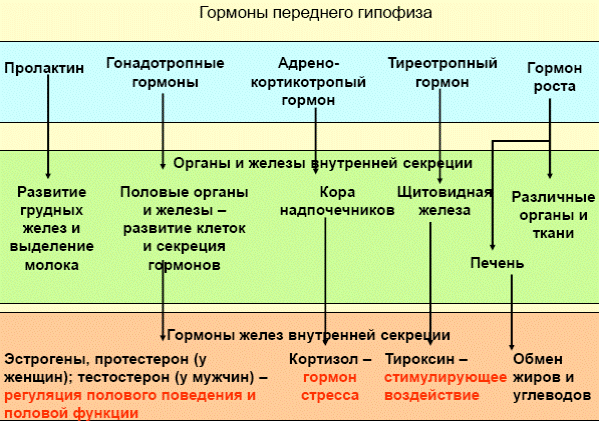
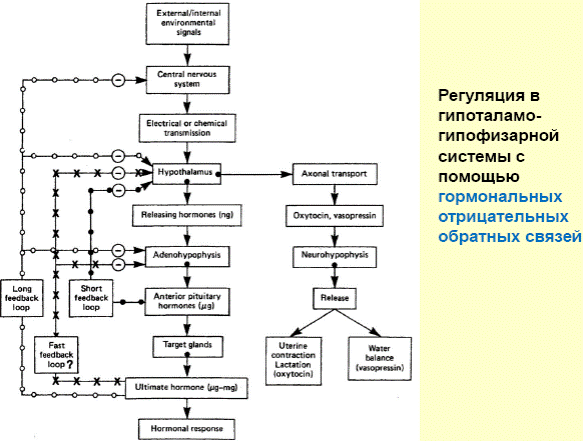
Гипоталамо-гипофизарная система: нейроэндокринные связи гипоталамуса с задним и передним гипофизом. Основные гормоны, выделяемые гипофизом, и их влияние на эндокринную систему и внутренние органы. Обратная связь в нейроэндокринной регуляции.
Генерация биоэлектрических потенциалов. В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала.
Все ткани организма могут находиться в двух состояниях:
1. состоянии относительного физиологического покоя;
2. состоянии активности, который наблюдается при раздражении ткани.
Существует 2 вида активного состояния тканей: возбуждение и торможение. Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани. Возбуждение характеризуется двумя группами признаков: неспецифическими и специфическими.
Неспецифические признаки возникают у всех возбудимых тканей вне зависимости от их строения:
изменение проницаемости клеточных мембран
изменение заряда клеточных мембран,
повышение потребления кислорода
повышение температуры
усиление обменных процессов
Специфические признаки различаются у различных тканей:
мышечная ткань – сокращение
железистая ткань – выделение секрета
нервная ткань – генерация нервного импульса.
Процесс возбуждения связан с наличием в мембране электрически (для ионов кальция и хлора) и химически (для ионов натрия и калия) управляемых каналов, которые могут открываться в ответ на соответствующее раздражение клетки.
Ионоселективные каналы. Для каждого из переносимых через мембрану вида ионов существуют самостоятельные транспортные системы – ионные каналы (натриевые, калиевые, кальциевые, каналы для хлора и т.д.). Ионный канал состоит из поры, воротного механизма, сенсора (индикатора) напряжения ионов в самой мембране и селективного фильтра.
Пора представляет собой молекулярное динамическое образование, которое может находиться в открытом и закрытом состоянии. Образована пора «транспортным» ферментом – белком с высокой каталитической активностью, который способен переносить ионы через мембрану со скоростью в 200 раз превышающей скорость простой диффузии.
Воротный механизм (ворота канала) расположен на внутренней стороне мембраны и представлен белковыми молекулами, способными к конформации (изменение пространственной конфигурации молекул). В тысячные доли секунды он открывает (активирует) и закрывает (инактивирует) канал и таким образом регулирует скорость передвижения ионов по нему и поступление их в цитоплазму.
Сенсор напряжения ионов в мембране представлен белковой молекулой, расположенной в самой мембране и способной реагировать на изменение мембранного потенциала.
Селективный фильтр находится в самом узком месте канала. Он определяет однонаправленное движение ионов через пору и ее избирательную проницаемость.
В развитии возбуждения выделяют 4 этапа:
1) предшествующее возбуждению состояние покоя (статическая поляризация);
2) деполяризацию;
3) реполяризацию
4) гиперполяризацию.
Деполяризация – сдвиг МП в сторону его уменьшения. Под действием раздражения открываются «быстрые» натриевые каналы, вследствие чего ионы Na лавинообразно поступают в клетку. Переход положительно заряженных ионов в клетку вызывает уменьшение положительного заряда на ее наружной поверхности и увеличение его в цитоплазме. В результате этого сокращается трансмембранная разность потенциалов, значение МП падает до 0, а затем по мере дальнейшего поступления Na в клетку происходят перезарядка мембраны и инверсия ее заряда (поверхность становится электроотрицательной по отношению к цитоплазме) – возникает потенциал действия (ПД). Электрографическим проявлением деполяризации является спайк, или пиковый потенциал.
Во время деполяризации, когда переносимый ионами Na положительный заряд достигает некоторого порогового значения, в сенсоре напряжения ионных каналов возникает ток смещения, который «захлопывает» ворота и «запирает» (инактивирует) канал, прекращая тем самым дальнейшее поступление Na в цитоплазму. Канал «закрыт» (инактивирован) вплоть до восстановления исходного уровня МП.
Реполяризация – восстановление исходного уровня МП. При этом ионы натрия перестают проникать в клетку, проницаемость мембраны для калия увеличивается, и он достаточно быстро выходит из нее. В результате заряд клеточной мембраны приближается к исходному. Электрографическим проявлением реполяризации является отрицательный следовой потенциал.
Гиперполяризация – увеличение уровня МП. Вслед за восстановлением исходного значения МП (реполяризация) происходит его кратковременное увеличение по сравнению с уровнем покоя, обусловленное повышением проницаемости калиевых каналов и каналов для Cl. В связи с этим поверхность мембраны приобретает избыточный по сравнению с нормой положительный заряд, а уровень МП становится несколько выше исходного. Электрографическим проявлением гиперполяризации является положительный следовой потенциал. На этом заканчивается одиночный цикл возбуждения.








