 2015-04-20
2015-04-20 4831
4831I. Основную группу клеток-продуцентов цитокинов в адаптивном иммунном ответе представляют лимфоциты. Покоящиеся клетки не секретируют цитокины. При распознавании антигена и при участии рецепторных взаимодействий (CD28-CD80/86 для Т-лимфоцитов и СD40-CD40L для В-лимфоцитов) происходит активация клеток, приводящая к транскрипции генов цитокинов, трансляции и секреции гликозилированных пептидов в межклеточное пространство.
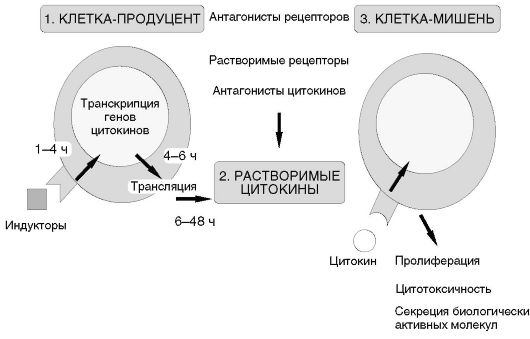 Рис. 7.1. Система цитокинов
Рис. 7.1. Система цитокинов
CD4 Т-хелперы представлены субпопуляциями: Тh0, Тh1, Тh2, Тh17, Tfh, которые различаются между собой спектром секретируемых цитокинов в ответ на различные антигены.
Тh0 вырабатывают широкий спектр цитокинов в очень низких концентрациях.
Направление дифференцировки Th0 определяет развитие двух форм иммунного ответа с преобладанием гуморальных или клеточных механизмов.
Природа антигена, его концентрация, локализация в клетке, тип антигенпрезентирующих клеток и определенный набор цитокинов регулируют направление дифференцировки Тh0.
Дендритные клетки после захвата и процессинга антигена представляют антигенные пептиды Th0 клеткам и вырабатывают цитокины, регулирующие направление их дифференцировки в эффекторные клетки. Роль индивидуальных цитокинов в данном процессе отражена на рис. 7.2. ИЛ-12 индуцирует синтез ИФНγ Т-лимфоцитами и ]ЧГК. ИФНу обеспечивает дифференцировку ТЫ1, которые начинают секретировать цитокины (ИЛ-2, ИФНу, ИЛ-3, ФНОа, лимфотоксины), регулирующие развитие реакций на внутриклеточные патогены
(гиперчувствительности замедленного типа (ГЗТ) и различные типы клеточной цитотоксичности).
ИЛ-4 обеспечивает дифференцировку Тh0 в Тh2. Активированные Тh2 вырабатывают цитокины (ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-13 и др.), определяющие пролиферацию В-лимфоцитов, их дальнейшую дифференцировку в плазматические клетки,и развитие реакций антителогенеза, преимущественно на внеклеточные патогены.
ИФНу негативно регулирует функцию Тh2-клеток и, наоборот, ИЛ-4, ИЛ-10, секретируемые Тh2, угнетают функцию Тh1 (рис. 7.3). Молекулярный механизм этой регуляции связан с транскрипционными факторами. Экспрессия Т-bet и STAT4, детерминированная ИФНу, направляет дифференцировку Т-клеток по пути Тh1 и супрессирует развитие Тh2. ИЛ-4 индуцирует экспрессию GATA-3 и STAT6, что соответственно обеспечивает превращение наивных ТЫ0 в Тh2-клетки (рис. 7.2).
В последние годы описана особая субпопуляция Т-клеток хелперов (Тh17), продуцирующих ИЛ-17. Члены семейства ИЛ-17 могут экспрессироваться активированными клетками памяти (CD4CD45RO), у5Т-клетками, NKT клетками, нейтрофилами, моноцитами под влиянием ИЛ-23, ИЛ-6, ТФРβ, вырабатываемых макрофагами и дендритными клетками. Основным дифференцировочным фактором у человека является ROR-C, у мышей - ROR-γ l Показана кардинальная роль ИЛ-17 в развитии хронического воспаления и аутоиммунной патологии (см. рис. 7.2).
Кроме того, Т-лимфоциты в тимусе могут дифференцироваться в естественные клетки-регуляторы (Treg), экспрессирующие поверхностные маркеры CD4+ CD25+ и транскрипционный фактор FOXP3. Эти клетки способны подавлять иммунный ответ, опосредуемый Тh1 и Тh2-клетками, путем прямого межклеточного контакта и синтеза ТФРβ и ИЛ-10.
Схемы дифференцировки клонов Тh0 и секретируемых ими цитокинов представлены на рис. 7.2 и 7.3 (см. также цв. вклейку).
Т-цитотоксические клетки (CD8+), естественные киллеры - слабые продуценты цитокинов, таких, как интерфероны, ФНОа и лимфотоксины.
Избыточная активация одной из субпопуляций Тh может определить развитие одного из вариантов иммунного ответа. Хроническая несбалансированность активации Тh способна привести к формированию иммунопатологических состояний, связанных с проявления-
ми аллергии, аутоиммунной патологии, хронических воспалительных процессов и др.
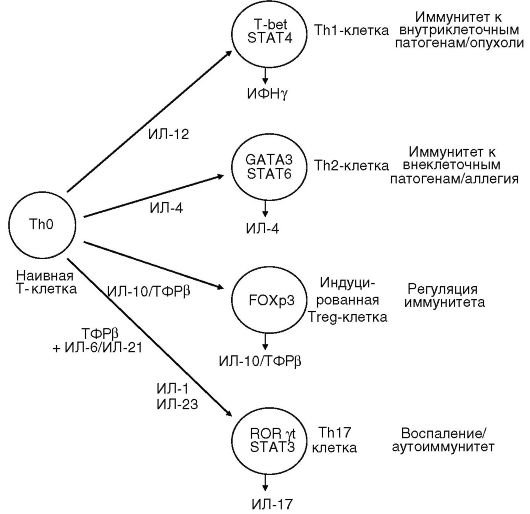 Рис. 7.2. Различные субпопуляции Т-лимфоцитов, продуцирующие цитокины
Рис. 7.2. Различные субпопуляции Т-лимфоцитов, продуцирующие цитокины
II. В системе врожденного иммунитета основными продуцентами цитокинов являются клетки миелоидного ряда. С помощью Toll-по- добных рецепторов (TLRs) они распознают сходные молекулярные структуры различных патогенов, так называемые патогенассоциированные молекулярные патерны (РАМП), например липополисахарид (ЛПС) грамотрицательных бактерий, липотейхоевые кислоты, пептидогликаны грамположительных микроорганизмов, флагеллин, ДНК, богатую неметилированными СрG повторами, и др. В результате
такого взаимодействия с TLR запускается внутриклеточный каскад передачи сигнала, приводящий к экспрессии генов двух основных групп цитокинов: провоспалительных и ИФН типа 1 (рис. 7.4, см. также цв. вклейку). Главным образом эти цитокины (ИЛ-1, -6, -8, -12, ФНОа, ГМ-КСФ, ИФН, хемокины и др.) индуцируют развитие воспаления и участвуют в защите организма от бактериальных и вирусных инфекций.
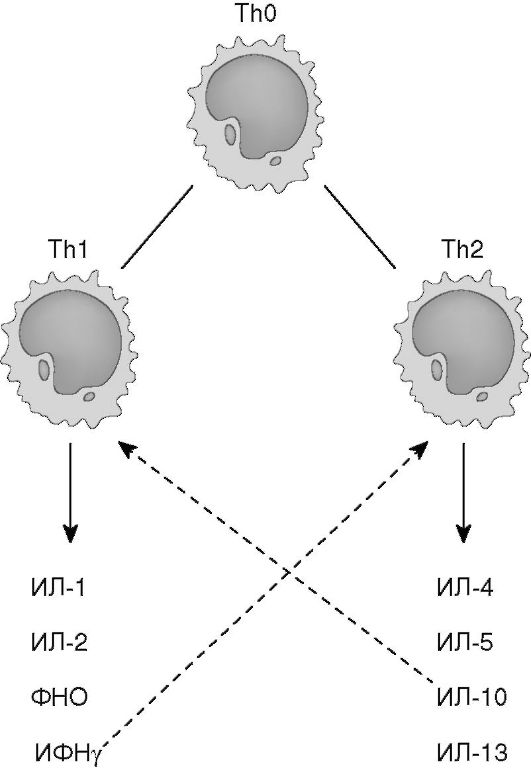 Рис. 7.3. Спектр цитокинов, секретируемых ТЫ1- и ТЫ2-клетками
Рис. 7.3. Спектр цитокинов, секретируемых ТЫ1- и ТЫ2-клетками
III. Клетки, не относящиеся к иммунной системе (клетки соединительной ткани, эпителия, эндотелия), конститутивно секретируют аутокринные факторы роста (ФРФ, ЕФР, ТФРр и др.). и цитокины, поддерживающие пролиферацию гемопоэтических клеток.
Цитокины и их антагонисты подробно описаны в ряде монографий (Ковальчук Л.В. и соавт., 2000; Кетлинский С.А., Симбирцев А.С.,
2008).
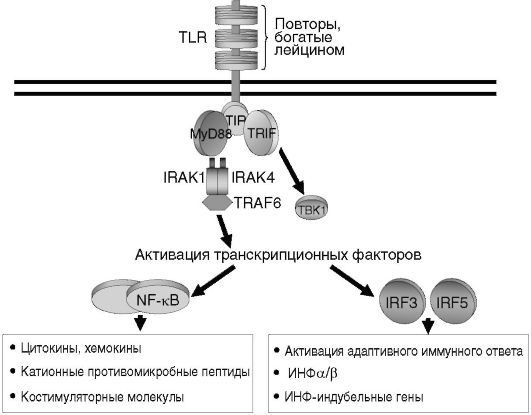 Рис. 7.4. TLR-опосредованная индукция выработки цитокинов клетками врожденного иммунитета
Рис. 7.4. TLR-опосредованная индукция выработки цитокинов клетками врожденного иммунитета
Избыточная экспрессия цитокинов небезопасна для организма и может привести к развитию чрезмерной воспалительной реакции, острофазового ответа. В регуляции выработки провоспалительных цитокинов принимают участие различные ингибиторы. Так, описан ряд веществ, которые неспецифически связывают цитокин ИЛ-1 и препятствуют проявлению его биологического действия (а2-макроглобулин, С3-компонент комплемента, уромодулин). Специфическими ингибиторами ИЛ-1 могут быть растворимые рецепторы-ловушки, антитела и рецепторный антагонист ИЛ-1 (ИЛ-1RA). При развитии воспаления происходит усиление экспрессии гена ИЛ-1RA. Но и в норме этот антагонист присутствует в крови в высокой концентрации (до 1 нг/мл и более), блокируя действие эндогенного ИЛ-1.








