 2015-04-30
2015-04-30 4809
4809Брожение — это внутренний окислительно-восстановительный процесс, при котором акцептором электронов служит органическая молекула и суммарная степень окисления образующихся продуктов не отличается от степени окисления сбраживаемого вещества. С. П. Костычев выдвинул положение о генетической связи процессов брожения и дыхания. При этом он опирался на следующие факты:
1.У высших растений был найден весь набор ферментов, который катализирует отдельные этапы процесса брожения.
2. При временном попадании в условия анаэробиоза высшие растения определенное время существуют за счет энергии, выделяющейся в процессе брожения. Правда, поскольку процесс брожения энергетически значительно менее эффективен, в анаэробных условиях рост растений приостанавливается. Кроме того, продукты брожения, в частности спирт, ядовиты, и их накопление приводит к гибели растения.
3. При добавлении к клеткам факультативных анаэробов (дрожжи) полусброженных Сахаров интенсивность дыхания у них резко возрастает, следовательно, полусброженные продукты являются лучшим субстратом дыхания по сравнению с неизмененными сахарами.
В настоящее время общепризнано, что первые этапы (гликолиз) протекают одинаково при процессах как дыхания, так и брожения. Поворотным моментом является образование пировиноградной кислоты. В аэробных условиях пировиноградная кислота распадается до СО2 и воды в результате декарбоксилирования и цикла Кребса (дыхание), тогда как в анаэробных она преобразуется в различные органические соединения (брожение). Организм обладает способностью при изменении условий переключать процессы, прекращая брожение и усиливая дыхание и наоборот. Впервые в опытах Пастера было показано, что в присутствии кислорода процесс брожения у дрожжей тормозится и заменяется процессом дыхания. Одновременно резко сокращается распад глюкозы. Это явление оказалось характерным для всех факультативных анаэробных организмов, включая и высшие растения, и получило название эффекта Пастера. Сокращение расхода глюкозы в присутствии кислорода целесообразно, поскольку при дыхательном распаде выход энергии значительно выше, а следовательно, глюкоза используется более экономно. Однако осуществление разбираемого эффекта требует специальных механизмов, которые будут рассмотрены далее.
В зависимости от получаемого продукта различают разные типы брожения. При спиртовом брожении пировиноградная кислота, образовавшаяся в процессе гликолиза, декарбоксилируется с образованием уксусного альдегида при участии фермента пируватдекарбоксилазы, а затем восстанавливается до этилового спирта ферментом алкогольдегидрогеназой:
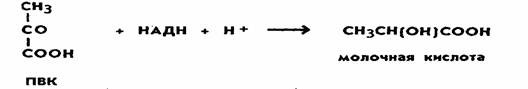
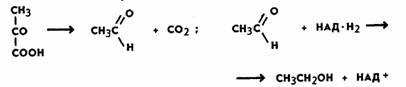 Ни та ни другая реакция не сопровождается образованием АТФ. В связи с этим выход АТФ при спиртовом брожении такой же, как при гликолизе (первой фазы брожения и дыхания), и составляет две молекулы при распаде 1 моль глюкозы. Восстановленные никотинамидные коферменты (НАД-Н2), образовавшиеся в процессе гликолиза, не поступают в дыхательную цепь (у анаэробных организмов ее и нет), а используются для восстановления уксусного альдегида до спирта. Следовательно, энергетический выход процессов брожения крайне низок. Разные микроорганизмы осуществляют и разные типы брожения. Так, молочнокислые бактерии накапливают молочную кислоту. При этом пировиноградная кислота восстанавливается до молочной кислоты:
Ни та ни другая реакция не сопровождается образованием АТФ. В связи с этим выход АТФ при спиртовом брожении такой же, как при гликолизе (первой фазы брожения и дыхания), и составляет две молекулы при распаде 1 моль глюкозы. Восстановленные никотинамидные коферменты (НАД-Н2), образовавшиеся в процессе гликолиза, не поступают в дыхательную цепь (у анаэробных организмов ее и нет), а используются для восстановления уксусного альдегида до спирта. Следовательно, энергетический выход процессов брожения крайне низок. Разные микроорганизмы осуществляют и разные типы брожения. Так, молочнокислые бактерии накапливают молочную кислоту. При этом пировиноградная кислота восстанавливается до молочной кислоты:
 Для некоторых оолигатных анаэробных микроорганизмов, например азотфиксирующей бактерии Clostridium pasteurianum, характерно образование в процессе брожения масляной кислоты (СН3СН2СН2СООН), СО2 и Н2. Процесс брожения служит источником энергии для облигатных (обязательных) или факультативных анаэробных организмов.
Для некоторых оолигатных анаэробных микроорганизмов, например азотфиксирующей бактерии Clostridium pasteurianum, характерно образование в процессе брожения масляной кислоты (СН3СН2СН2СООН), СО2 и Н2. Процесс брожения служит источником энергии для облигатных (обязательных) или факультативных анаэробных организмов.
Энергетический баланс процесса дыхания
Подводя итоги энергетики процесса дыхания, подсчитаем, сколько всего молекул АТФ может образоваться при распаде одной молекулы глюкозы. В первую анаэробную фазу дыхания при распаде одной молекулы глюкозы до двух молекул пировиноградной кислоты в процессе субстратного фосфорилирования накапливаются две молекулы АТФ. Одновременно на этой фазе дыхания при окислении ФГА до ФГК в цитозоле образуются две молекулы восстановленных коферментов (2 НАД-Н2). Они диффундируют через наружную мембрану и окисляются в дыхательной цепи благодаря наличию у растений НАД-Н-дегидрогеназы, локализованной на наружной поверхности внутренней мембраны. При этом синтезируется 6 молекул АТФ. В аэробной фазе дыхания при окислении пировиноградной кислоты образуются 4НАД-Н2. Их окисление в дыхательной цепи приводит к образованию 12 АТФ. Кроме того, в цикле Кребса восстанавливается одна молекула флавиновой дегидрогеназы (ФАД-Н2). Окисление этого соединения в дыхательной цепи приводит к образованию 2 АТФ, поскольку одно фосфорилирование не происходит. При окислении молекулы а-кетоглутаровой кислоты до янтарной энергия непосредственно накапливается в одной молекуле АТФ (субстратное фосфорилирование).
Таким образом, окисление одной молекулы пировиноградной кислоты сопровождается образованием ЗСО2 15 молекул АТФ. Однако при распаде молекулы глюкозы образовались две молекулы пировиноградной кислоты. Следовательно, всего в аэробной фазе дыхания образуется 6 молекул СО2 и 30 молекул АТФ плюс 8АТФ в анаэробной фазе. Итого 6 молекул СО2 и 38 молекул АТФ образуется в процессе окислительно-дыхательного распада молекулы гексозы. На образование 38 молекул АТФ затрачено 38-30,6 кДж = 1162,8 кДж. Всего при сжигании 1 моль глюкозы выделяется 2824 кДж: C6H12O6 + 6O2->6CO2 + 6H2O + 2824 кДж. Таким образом, КПД процесса дыхания при самых благоприятных условиях составляет около 40%.
Подводя итоги, можно сказать, что биологическое окисление — это многоступенчатый ферментативный процесс, сопровождаемый выделением энергии.
Промежуточный обмен веществ. Совокупность химических превращений веществ, которые происходят в организме, начиная с момента их поступления в кровь и до момента выделения конечных продуктов обмена из организма, называют промежуточным, или межуточным обменом (промежуточным метаболизмом). Промежуточный обмен может быть разделен на два процесса: катаболизм (диссимиляция) и анаболизм (ассимиляция). Катаболизмом называют ферментативное расщепление сравнительно крупных органических молекул, осуществляемое у высших организмов, как правило, окислительным путем. Катаболизм сопровождается освобождением энергии, заключенной в сложных структурах органических молекул, и запасанием ее в форме энергии фосфатных связей АТФ. Анаболизм представляет собой ферментативный синтез крупномолекулярных клеточных компонентов, таких, как полисахариды, нуклеиновые кислоты, белки, липиды, а также некоторых их биосинтетических предшественников из более простых соединений. Анаболические процессы происходят с потреблением энергии. Катаболизм и анаболизм происходят в клетках одновременно и неразрывно связаны друг с другом. По существу, их следует рассматривать не как два отдельных процесса, а как две стороны одного общего процесса — метаболизма, в котором превращения веществ теснейшим образом переплетены с превращениями энергии.
Подробный анализ метаболических путей показывает, что расщепление основных пищевых веществ в клетке представляет собой ряд последовательных ферментативных реакций, составляющих три главные стадии катаболизма. На первой стадии крупные органические молекулы распадаются на составляющие их специфические структурные блоки. Так, полисахариды расщепляются до гексоз или пентоз, белки — до аминокислот, нуклеиновые кислоты — до нуклеотидов и нуклеозидов, липиды — до жирных кислот, глицерина и других веществ. Все эти реакции протекают в основном гидролитическим путем (см. Гидролиз) и количество энергии, освобождающейся на этой стадии, очень невелико — менее 1%. На второй стадии катаболизма формируются еще более простые молекулы, причем число их типов существенно уменьшается. Очень важно, что на второй стадии образуются продукты, которые являются общими для обмена разных веществ. Эти продукты — ключевые соединения, представляющие собой как бы узлы, соединяющие разные пути метаболизма. К таким соединениям относятся, например, пируват (пировиноградная кислота), образующийся при распаде углеводов, липидов и многих аминокислот; ацетил-КоА; объединяющий катаболизм жирных кислот, углеводов и аминокислот; a-кетоглутаровая кислота, оксалоацетат (щавелевоуксусная кислота), фумарат (фумаровая кислота) и сукцинат (янтарная кислота), образующиеся из разных аминокислот, и др. Продукты, полученные на второй стадии катаболизма, вступают в третью стадию катаболизма, которая известна как цикл трикарбоновых кислот (терминальное окисление, цикл лимонной кислоты, цикл Кребса). В ходе этой стадии все продукты в конечном счете окисляются до СО2 и воды. Практически почти вся энергия освобождается на второй и третьей стадиях катаболизма.
Процесс анаболизма тоже проходит три стадии. Исходными веществами для него служат те продукты, которые подвергаются превращениям на третьей стадии катаболизма. Т.о., третья стадия катаболизма является в то же время первой, исходной стадией анаболизма. Реакции, протекающие на этой стадии, выполняют как бы двойную функцию. С одной стороны, они участвуют в завершающих этапах катаболизма, а с другой — служат и для анаболических процессов, поставляя вещества-предшественники для последующих стадий анаболизма. Нередко такие реакции называют амфиболическими. На этой стадии, например, начинается синтез белка. Исходными реакциями этого процесса можно считать образование некоторых a-кетокислот. На следующей, второй стадии в ходе реакций аминирования или трансаминирования эти кетокислоты превращаются в аминокислоты, которые на третьей стадии анаболизма объединяются в полипептидные цепи. В результате ряда последовательных реакций происходит также синтез нуклеиновых кислот, липидов и полисахаридов. Лишь в 60—70 гг. 20 в. выяснилось, что пути анаболизма не являются простым обращением процессов катаболизма. Это связано с энергетическими особенностями химических реакций. Некоторые реакции катаболизма практически необратимы, т.к. их протеканию в обратном направлении препятствуют непреодолимые энергетические барьеры. В ходе эволюции были выработаны другие, обходные реакции, сопряженные с затратой энергии макроэргических соединений.
Катаболический и анаболический пути отличаются, как правило, и локализацией в клетке. Например, окисление жирных кислот ацетата осуществляется с помощью набора митохондриальных ферментов, тогда как синтез жирных кислот катализирует другая система ферментов, находящихся в цитозоле. Именно благодаря разной локализации катаболические и анаболические процессы в клетке могут протекать одновременно.
Т.о., пути метаболизма чрезвычайно многообразны. Однако в этом многообразии можно усмотреть проявление удивительного единства, которое является наиболее типичной и специфической чертой обмена веществ. Это единство состоит в том, что от бактерий до самой высокодифференцированной ткани высшего организма биохимической реакции не только внешне сходны, например по балансовым уравнениям и внешним эффектам, но и абсолютно тождественны во всех деталях. Другим проявлением такого единства следует считать наблюдаемое также на всем пути эволюции циклическое протекание важнейших метаболических процессов, например цикл трикарбоновых кислот, цикл мочевины, пентозный путь и др. Видимо, и сами биохимические реакции, отобранные и закрепленные в ходе эволюции, и цикличность их протекания оказались оптимальными для обеспечения физиологических функций организма.

