2015-04-30
2015-04-30 13068
13068Функции РНК различаются в зависимости от вида рибонуклеиновый кислоты.
1) Информационная РНК (и-РНК).
2) Рибосомная РНК (р-РНК).
3) Транспортная РНК (т-РНК).
4) Минорные (малые) РНК. Это молекулы РНК, чаще всего с небольшой молекулярной массой, располагающиеся в различных участках клетки (мембране, цитоплазме, органеллах, ядре и т.д.). Их роль до конца не изучена. Доказано, что они могут помогать созреванию рибосомной РНК, участвуют в переносе белков через мембрану клетки, способствуют редупликации молекул ДНК и т.д.
5) Рибозимы. Недавно выявленный вид РНК, принимающие активное участие в ферментативных процессах клетки в качестве фермента (катализатора).
6) Вирусные РНК. Любой вирус может содержать только один вид нуклеиновой кислоты: либо ДНК либо РНК. Соответственно, вирусы, имеющие в своём составе молекулу РНК, получили название РНК-содержащие. При попадании в клетку вируса данного типа может происходить процесс обратной транскрипции (образование новых ДНК на базе РНК), и уже вновь образовавшаяся ДНК вируса встраивается в геном клетки и обеспечивает существование, а также размножение возбудителя. Вторым вариантом сценария является образование комплиментарной РНК на матрице поступившей вирусной РНК. В этом случае, образование новых вирусных белков, жизнедеятельность и размножение вируса происходит без участия дезоксирибонуклеиновой кислоты только на основании генетической информации, записанной на вирусной-РНК. Рибонуклеиновые кислоты. РНК, строение, структуры, виды, роль. Генетический код. Механизмы передачи генетической информации. Репликация. Транскрипция
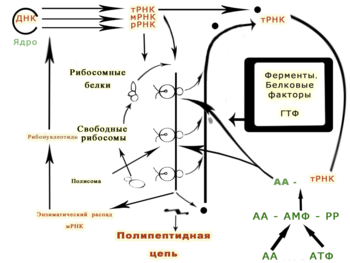
Рибосомная РНК.
На долю рРНК приходится 90% всей РНК клетки, она характеризуется метаболической стабильностью. У прокариот различают три различных типа рРНК с коэффициентами седиментации 23S,16S и 5S; у эукариот четыре типа:-28S, 18S,5S и 5,8S.
РНК этого типа локализованы в рибосомах и участвуют в специфическом взаимодействии с рибосомными белками.
Рибосомные РНК имеют форму вторичной структуры в виде которых двуспиральных участков, соединенных изогнутой одиночной цепью. Белки рибосомы связаны преимущественно с однотяжевыми участками молекулы.
Для рРНК характерно наличие модифицированных оснований, однако в значительно меньшем количестве, чем в тРНК. В рРНК присутствуют главным образом метилизированные нуклеотиды, причем метильные группы присоединены либо к основанию, либо к 2/- OH- группе рибозы.
Транспортная РНК.
Молекулы тРНК представляют собой единую цепь, состоящую из 70-90 нуклеотидов, с молекулярной массой 23000-28000 и константой седиментации 4S. В клеточной РНК транспортная РНК составляет 10-20%. Молекулы тРНК обладают способностью ковалентно связываться с определенной аминокислотой и соединяться через систему водородных связей с одним из нуклеотидных триплетов молекулы мРНК. Таким образом, тРНК реализуют кодовое соответствие между аминокислотой и отвечающим ей кодоном мРНК. Для выполнения адапторной функции тРНК должны иметь вполне определенную вторичную и третичную структуру.
Каждая молекула тРНК обладает постоянной вторичной структурой, имеет форму двумерного клеверного листа и состоит из спиральных участков, образованных нуклеотидами одной и той же цепи, и расположенных между ними одноцепочечных петель. Количество спиральных областей достигает половины молекулы.Неспаренные последовательности образуют характерные структурные элементы (ветви),имеющие типичные ветви:
А) акцепторный стебель, на 3/-OH конце которого в большинстве случаев расположен триплет ЦЦА. К карбоксильной группе концевого аденозина с помощью специфического фермента присоединяется соответствующая аминокислота;
Б) псевдоуридиновая или Т Ц-петля, состоит из семи нуклеотидов с обязательной последовательностью 5/-Т ЦГ-3/, в которой содержится псевдоуридин; предполагается что Т Ц-петля используется для связывания тРНК с рибосомой;
В) дополнительная петля-различная по размеру и составу в разных тРНК;
Г) антикодоновая петля состоит из семи нуклеотидов и содержит группу из трех оснований (антикодон), которая комплементарна триплету (кодону) в молекуле иРНК;
Д) дигидроуридиловая петля (D-петля), состоящая из 8-12 нуклеотидов и содержащая от одного до четырех дигидроуридиловых остатков;считается, что D-петля используется для связывания тРНК со специфическим ферментом (аминоацил-тРНК-синтетаза).
Третичная укладка молекул тРНК является весьма компактной и имеет Г-образную форму. Угол подобной структуры образован дигидроуридиновым остатком и Т Ц-петлей, длинное колено образует акцепторный стебель и Т Ц-петля, а короткое-D-петля и антикодоновая петля.
В стабилизации третичной структуры тРНК участвуют поливалентные катионы (Mg2+, полиамины), а также водородные связи между основаниями и фосфодиэфирным остовом.
Сложная постранственная укладка молекулы тРНК обусловлена множественными высокоспецифичными взаимодействиями как с белками, так и с другими нуклеиновыми кислотами (рРНК).
Транспортная РНК отличается от других типов РНК высоким содержанием минорных оснований-в среднем 10-12 оснований на молекулу, однако общее число их а тРНК растет по мере продважения организмов по эволюционной лестнице. В тРНК выявлены различные метилированные пуриновые (аденин, гуанин) и пиримидиновые (5-метилцитозин и рибозилтимин) основания, серосодержащие основания (6-тиоурацил), но наиболее распростран(6-тиоурацил), но наиболее распространенным минорным компонентом является псевдоуридин. Роль необычных нуклеотидов в молекулах тРНК пока не ясна, однако пологают, что чем ниже уровень митилирования тРНК, тем она менее активна и специфична.
Локализация модифицированных нуклеотидов строго фиксирована. Наличие минорных оснований в составе тРНК обуславливает устойчивасть молекул к действию нуклеаз и, кроме того, они участвуют в поддержании определенной структуры, так как подобные основания не способны к нормальному спариванию и препятствуют образованию двойной спирали. Таким образом, наличие модифицированных оснований в составе тРНК обуславливает не только её структуру, но также и многие специальные функции молекулы тРНК.
В большинстве клеток эукариот содержится набор различных тРНК. Для каждой аминокислоты имеется не менее чем по одной специфической тРНК. тРНК связывающие одну и ту же аминокислоту, называют изоакцепторными. Каждый тип клеток в организме отличется своим соотношением изоакцепторных тРНК.
Матричная (информационная)
Матричная РНК содержит генетическую информацию о последовательности аминокислот для основных ферментов и других белков, т.е. служит матицей для биосинтеза полипептидных цепей. На долю мРНК в клетке приходится 5% от общего количества РНК. В отличий от рРНК и тРНК,мРНК гетерогенна по размерам,её молекулярная масса находится в пределах от 25 103 до 1 106; мРНК характеризуется широким диапазоном констант седиментации (6-25S). Наличие в клетке цепи мРНК переменной длинны отражает разнообразие молекулярных масс белков, синтез которых они обеспечивают.
По своему нуклеотидному составу мРНК соответствует ДНК из той же клетки,т.е. является комплементарной к одной из цепи ДНК. В последовательности нуклеотидов (первичная структура) мРНК заложена информация не только о структуре белка, но и о вторичной структуре самих молекул мРНК. Вторичная структура мРНК формируется за счет взаимокомплементарных последовательностей, содержание которых у РНК различного происхождения сходно и состовляет от 40 до 50%. Значительное количество спаренных участков может образовываться в 3/ и 5/-зонах мРНК.
Анализ 5/-концов областей 18s рРНК показал,что в них имеются взаимокомплементарные последовательности.
Третичная структура мРНК формируется главным образом за счет водородных связей, гидрофобного взаимодействия, геометрического и стерического ограничения, электрических сил.
Матричная РНК представляет собой метаболически активную и относительно не стабильную, короткоживущую форму. Так, мРНК микроорганизмов характеризуется бысрым обновлением, ивремя жизни её состовляет несколько минут. Вместе с тем для организмов, клетки которых содержат истинные ограниченые мембраной ядра, продолжительность жизни мРНК может достигать многих часов и даже несколько дней.
Стабильность мРНК может определяться различного рода модификациями её молекулы. Так, обнаружено, что 5/-концевая последовательность мРНК вирусов и эукариот метилирована,или «заблокирована». Первым нуклеотидом в 5/-терминальной структуре кэпа является 7-метилгуанин, который связан со следующим нуклеотидом 5/-5/-пирофосфатной связью. Второй нуклеотид метилирован по C-2/-рибозного остатка, а в третьем нуклеотиде метильной группы может и не быть.
Ещё одной способностью мРНК является то, что на 3/-концах многих молекул мРНК эукариотических клеток имеются относительно длинные последовательности адениловых нуклеотидов, которые присоединяются к молекулам мРНК с помощью специальных ферентов уже после завершения синтеза. Реакция протекает в клеточном ядре и цитоплазме.
На 3/- и 5/- концах мРНК модифицируемые последовательности составляют около 25% от общей длины молекулы. Считают, что 5 /– кэпы и 3/-поли-А – последовательности необходимы либо для стабилизации мРНК, предохраняющей её от действия нуклеаз, либо для регулирования процесса трансляции.
РНК-интерференция
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов.
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов.
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5' и 3' нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте - особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
Как и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на:
содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
«минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина.
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны.