| Визуали- зация | а) В процессе деления клетки (митоза или мейоза) хромосомы резко конденсируются, в результате чего значительно  укорачиваются и утолщаются. б) После этого они становятся видимыми даже на светооптическом уровне. укорачиваются и утолщаются. б) После этого они становятся видимыми даже на светооптическом уровне. |
| Различие формы | При конденсации разные хромосомы приобретают разную форму и по этому признаку могут быть идентифицированы. |
| Опреде- ление | а) Морфологию хромосом, как правило, описывают по их состоянию на стадии метафазы или анафазы митоза. б) Соответственно, хромосомы на этих стадиях называются метафазными и анафазными. |
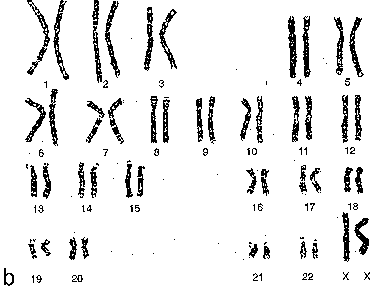
| Гомология хромосом | а) Изучение этих хромосом, в частности, показывает, что хромосомы попарно гомологичны друг другу, а состав одной (23-й) пары хромосом зависит от пола человека. б) Очевидно, это относится и к интерфазным хромосомам. |
| Двойная спираль | Любая из молекул хромосомной (ядерной) ДНК – очень протяжённая линейная структура, состоящая из двух полинуклеотидных цепей, которые являются взаимно комплементарными и закручены в двойную спираль. |
| Компле- ментар- ность | а) Комплементарность означает, что против аденилового нуклеотида одной цепи всегда стоит тимидиловый нуклеотид другой цепи, а против гуанилового нуклеотида всегда стоит цитидиловый. б) В этих пaрах (А-Т и Г-Ц) нуклеотиды как бы дополняют друг друга по своей пространственной конфигурации, что обеспечивает максимально плотную упаковку цепей в двойной спирали. |
| Длина | а) Молекулы хромосомных ДНК существенно различаются по количеству в них нуклеотидных пар (н.п.) и, соответственно, по своей длине. б) Поэтому различаются по длине и хромосомы – как интерфазные, так и метафазные. в) Средняя же длина одной молекулы ДНК человека – 4 см (120.000.000 н.п). г) Следовательно, общая протяжённость всех 46 молекул ДНК, находящихся в хромосомах одной клетки, – примерно 185 см. |
| Объясне- ние высокой длины ДНК | Такая огромная (для молекулярного уровня) длина обусловлена тем, что совокупность внутриядерных молекул ДНК (а значит, и ядро каждой клетки) содержит информацию обо всём организме человека, а именно о развитии, структуре и функционировании всех его систем и органов. |
| Информа- ция в ДНК | Конкретно, в наборе молекул ядерной ДНК каждой клетки содержится следующая информация: о первичной структуре (последовательности аминокислот) всех белков организма (исключение – некоторые белки митохондрий, кодируемые митохондриальной ДНК; п. 3.3.2.2), о первичной структуре (последовательности нуклеотидов) примерно 60 видов транспортных РНК (тРНК) и 4 видов рибосомных РНК(рРНК), а также, видимо, о программе использования данной информации в разных клетках в разные моменты онтогенеза. |
| Этапы передачи инфор- мации | Передача информации о структуре белка включает 3 этапа: транскрипцию ДНК – образование на участке ДНК как на матрице матричной РНК (мРНК), а точнее, её предшественника (пре-мРНК); созревание мРНК (процессинг) и перемещение её в цитоплазму. трансляцию мРНК(п. 3.3.1.3.) – синтез на рибосомах (в цитоплазме) полипептидной цепи (т.е. последовательности аминокислот) в соответствии с последовательностью нуклеотидных триплетов (кодонов) в мРНК. |
| Строение РНК | Напомним: РНК любого вида (мРНК, рРНК, тРНК) представляет собой одиночную цепь, состояшую из рибонуклеотидов – адениловых, уридиловых, гуаниловых и цитидиловых. |
| Основные процессы | В рамках реализации данной функции в ядрах происходят указанные ранее процессы: 1. транскрипцияопределённых участков ДНК (синтез пре-мРНК), 2. созревание (процессинг) мРНК, 3. синтез и созревание тРНК и рРНК. |
| Суть процес- синга | а) Пре-мРНК обычно содержат не только кодирующие последовательности нуклеотидов – экзоны, но и некодирующие – интроны (чередующиеся с экзонами). б) Поэтому одно из событий созревания пре-мРНК – это т.н. сплайсинг: вырезание интронов и сшивание экзонов в единую цепь мРНК. в) Затем к концам этой цепи присоединяются дополнительные последовательности, которые необходимы для связывания с рибосомами и для защиты мРНК от преждевременного разрушения экзонуклеазами. |
| Другие процессы в ядре (той же направ- ленности) | К прочтению информации ДНК имеют отношение также следующие внутриядерные процессы: 4.формирование субъединиц рибосом (из рРНК и поступающих из цитоплазмы рибосомных белков) и 5. разнообразныерегуляторные процессы(в том числе модификация ДНК и хромосомных белков), изменяющие активность разных участков ДНК. |
| 1. Репарация ДНК | а) Репарация ДНК – это обнаружение и исправление возникающих повреждений структуры ДНК. б) Репарация совершается практически постоянно, но её эффективность меньше 100% и к тому же у взрослых людей постепенно снижается – примерно на 1% за год. |
| 2. Реплика- ция ДНК | а)Репликация(удвоение) ДНК происходит перед любым делением клетки (кроме второго деления мейоза). б) В продуктах репликации – дочерних молекулах ДНК одна из цепей оказывается старой, а вторая – новой (синтезированной на первой по принципу комплементарности). Это означает, что репликация является полуконсервативным процессом. |
| 3. Конден- сация хромосом | Конденсация хромосом осуществляется уже в ходе деления клетки, в его начальной фазе, и облегчает последующее расхождение хромосом к полюсам делящейся клетки. |
| 1-2. Репарация и апоптоз | а) В указанных клетках значительно ужесточаются (по сравнению с соматическими клетками) требования к качеству наследственного материала, передаваемого потомству. б) Поэтому, как считают, резко повышаются интенсивность репарационных процессов и вероятность апоптоза. |
| 3. Конден- сация хромосом | а) При созревании сперматозоидов происходит гораздо более значительное уплотнение хромосом, чем при делениях клеток. б) Такая сверхконденсация хромосом достигается путём смены хромосомных белков. |
| 4-5. Конъюга- ция и кросс- синговер | а) Кроме того, добавляются новые процессы – конъюгация и кроссинговер. б) Они совершаются в ходе особого деления – мейоза – и заключаются в том, что гомологичные хромосомы вначале прилегают друг к другу (это обозначается термином “конъюгация”), а затем обмениваются некоторыми своими участками (кроссинговер). в) Данный обмен повышает разнообразие генетического материала в половых клетках. |
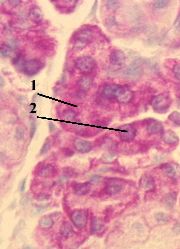
| 2. Препарат - включение Н3–уридина в РНК. Окраска гематоксилин-эозином. |
| 1. а) На представленном снимке мы видим, что меченое вещество (2) сосредоточено, главным образом, в ядрах (1) клеток. б) Это и отражает тот факт, что в ядрах происходит синтез всех видов РНК - мРНК, тРНК и рРНК. | 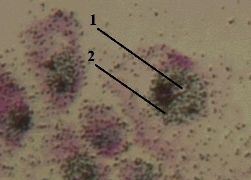 Полный размер Полный размер |
| 2. Наличие метки в других частях препарата объясняется, например, тем, что какая-то часть меченого вещества (Н3–уридина) не успела включиться в состав РНК, а какая-то часть новообразованной РНК, наоборот, уже успела выйти из ядра в цитоплазму. |
| 2. Природа эу- и гетеро- хроматина | а) Из предыдущего изложения ясно, что эухроматин – это те участки хромосом или целые хромосомы, которые находятся в деконденсированном (диффузном) состоянии, а гетерохроматин– это, напротив, конденсированные (и потому электроноплотные) фрагменты хромосом или целые хромосомы. б) В связи с этим, эухроматин ещё называют диффузным хроматином, а гетерохроматин – конденсированным хроматином. |
| 3. Световой уровень | а) При световой микроскопииэухроматин, как уже отмечалось, в ядре не выявляется. б) Те же глыбки, которые обычно обозначаются как “глыбки хроматина”, представляют собой на самом деле лишь гетерохроматин. |
| 4. Актив- ность эу- и гетеро- хроматина | а) В конденсированных участках хромосом ДНК недоступна для разнообразных внутриядерных ферментов – в том числе для ферментного комплекса, осуществляющего транскрипцию ДНК. б) В связи с этим, гетерохроматин функционально неактивен, а эухроматин, наоборот,активно участвует в транскрипции ДНК (синтезе пре-мРНК и других РНК). |
| Половые хромосомы у мужчин | а) У мужчин в наборе хромосом каждой клетки содержится, как известно, по одной Х- и Y-половой хромосоме. б) Обе они находятся в деконденсированном состоянии, т.е. входят во фракцию эухроматина. |
| Половые хромосомы у женщин | а)У женщин в клетках содержатся по две Х-хромосомы. б) А. Одна из них деконденсирована. Б. Вторая же Х-хромосома всегда находится в конденсированном состоянии, образуя в ядре компактное тельце - половой хроматин (тельце Барра). |
| 4. Препарат - половой хроматин в лейкоцитах (мазок крови женщины). Окраска азур 2-эозином. |
| Обнаруже- ние полового хроматина в мазке крови | а) Для обнаружения полового хроматина обычно исследуют мазок крови. б) В нейтрофильных лейкоцитах женщин половой хроматин выявляется | 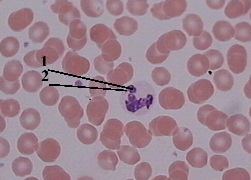 Полный размер Полный размер |
| в виде барабанной палочки (2), находящейся в одном из сегментов ядра (1). в) По этому признаку в судебной медицине отличают кровь женщин от крови мужчин. |
| Другой способ | Другой часто используемый в тех же целях объект – эпителиальные клетки слизистой оболочки полости рта. |
| Перечень компо- нентов | Как уже отмечалось, каждая из 46 хромосом представляет собой дезоксирибонуклеопротеин (ДНП), т.е. комплекс очень длинной молекулы ДНК (или двух таких молекул – после репликации ДНК) с определёнными белками, а также с небольшим количеством РНК. |
| РНК хромосом | а) Что касается РНК, то это либо незавершённые продукты транскрипции, либо молекулы, выполняющие регуляторные, структурные или иные функции. б) В последнее время особенно подчёркивается регуляторное влияние (на активность генов) т.н. малых молекул РНК. |
| Белки | Содержание же белков в хромосомах (и в хроматине в целом) намного выше содержания не только РНК, но и ДНК: по сравнению с последним (содержанием ДНК) оно обычно в 1,3–1,7 раза больше (по массе). |
| Виды гистонов | а) Основные белки представлены гистонами пяти видов: Н1, Н2А, Н2В, Н3 и Н4. б) Всего на них приходится 60-80% массы всех хромосомных белков. |
| Особен- ности состава | Гистоны обогащены аминокислотами с положительно заряженными радикалами (аргинин, лизин) и аминокислотами с гидрофобными радикалами (валин и т.п.). |
| Свойства | При этом благодаря радикалам аргинина и лизина гистоны взаимодействуют с ДНК (где каждый нуклеотид содержит отрицательно заряженную фосфатную группу), а благодаря гидрофобным радикалам – друг с другом. |
| Функция | а) Указанные взаимодействия приводят к образованию первого (нуклеосомного) и второго (нуклеомерного) уровней укладки ДНК в хромосоме (о чём будет сказано ниже). б) Таким образом, гистоны выполняют важную структурообразующую функцию. |
| Кислые белки чрезвычайно разнообразны: имеется, по крайней мере, несколько сотен их различных представителей. |
| Структур- ные белки | По всей видимости, некоторые кислые белки тоже играют структурную роль, участвуя в образовании высших (наднуклеомерных) уровней укладки хромосом. |
| Ферменты | а) Другую группу составляют многочисленные ферменты, обеспечивающие процессы репликации, модификации, репарации, транскрипции и даже разрушения ДНК. б) Это не означает, что все работающие на ДНК ферменты постоянно связаны с ней: какие-то ферменты могут находиться в кариоплазме. |
| Регуля- торные белки | а) Наконец, самой разнообразной по составу, видимо, является группа регуляторных белков. б) Эти белки контролируют активность вышеуказанных ферментов, а также доступность тех или иных участков ДНК для этих ферментов. |
| Кол-во нуклеосом | Молекула ДНК участвует в образовании очень большого числа нуклеосом (в среднем, 600.000). |
| ДНК - цепь "бусин" | В результате, на данном уровне организации каждая хромосома представляет собой длинную нить "бусинок"-нуклеосом толщиной 10 нм, а деконденсированный хроматин имеет мелкогранулярную структуру (2). | 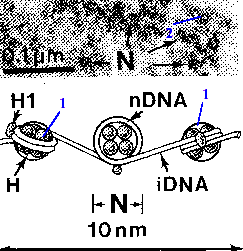 |
| Влияние нуклеосом на доступ- ность ДНК | а) Нуклеосомная организация не препятствует действию на ДНК внутриядерных ферментов. б) Хотя не исключено, что в момент прохождения ферментного комплекса по какому-либо участку ДНК последний на короткое время освобождается от связи с гистоновыми октамерами. Но затем эта связь быстро восстанавливается. |
| Новообра- зуемые цепи ДНК | Новообразуемые участки строящейся цепи ДНК (при репликации ДНК) тоже почти сразу приобретают нуклеосомную структуру. |
| Итоги | а) Таким образом, нуклеосомный уровень укладки присущ хромосомам практически постоянно. б)Заметим также: по сравнению с молекулой ДНК, нуклеосомная нить оказывается в 6,2 раза короче – благодаря закручиванию ДНК вокруг каждого октамера. |
| Роль гистона Н1 | Данный уровень образуется (при определённых условиях) в результате взаимодействия друг с другом молекул гистона Н1, находящихся в составе нуклеосомной нити. |
| Названия структуры | При этом нуклеосомная нить конденсируется в более плотную структуру толщиной около 30 нм: нуклеомерную нить,или хроматиновую фибриллу. |
| Описание структуры | а) Полагают, что нуклеомерная нить представляет собой суперспираль, причём такую, в которой вновь (как в нуклеосомной цепи) чередуются глобулярные и линкерные участки. б) Иначе говоря, это цепочка “супербус” (нуклеомеров), где в одной “супербусине” – 5-8 нуклеосом. |
| Блокиро- вание активности генов | а) Однако в этой цепочке ДНК уже недоступна для ферментных комплексов (в т.ч. того, который осуществляет транскрипцию). б) Поэтому данный уровень организации отсутствует у эухроматина. |
| Сокраще- ние длины хромосомы | Там же, где нуклеомерная структура имеется, длина хромосомы (или её отдела, если конденсируется только он) уменьшается ещё примерно в 6-7 раз. |
| Факторы и принцип укладки | а) Дальнейшая компактизация хромосом происходит под влиянием уже не гистонов, а определённых кислыхбелков. б) И начинает использоваться другой принцип укладки: образование петель, или складок. |
| Описание структуры | а) Так, хроматиновая нить образует множество петель, а последние собираются в розетки. б) Каждую розетку составляет несколько соседних петель, которые прикрепляются своими основаниями к общему белковому центру. в) Подобные центры в большом количестве содержатся в ядерном матриксе. |
| Названия элементов структуры | Таким образом, на этом уровне организации хромосома представляет собой цепь, состоящую из розеток, которые называются также петельными доменами и хромомерами. |
| Особен- ность гетеро- хроматина | В гетерохроматине розетки, видимо, расположены более-менее рыхло, т.е. дальше компактизация хромосом не идёт. |
| Вероятный способ организа- ции данного уровня | а) Хромонема спирализуется или тоже складывается в петли. б) При этом петли, возможно, образуют группы, которые составляют сегмент хроматиды и связаны друг с другом короткими линкерными участками. в) Это соответствует тому обстоятельству, что под действием повреждающих факторов хромосома может распадаться на сегменты. |
| Название структуры | Продукт данного (и последнего) этапа конденсации – хроматида толщиной 700 нм, одна из двух “половинок” метафазной хромосомы. |
| Параметры хромосомы | а)Толщина всей хромосомы в состоянии максимальной компактизации – вдвое больше: 1400 нм б) Зато длина хромосомы человека за счёт всех уровней укладки сокращается, в итоге, в 10.000 раз: молекулы ДНК общей длиной ~200 см укладываются в метафазных хромосомах общей длиной ~200 мкм. |
| Уровень укладки | Пояснение | Толщина | Сокращение длины |
| I. Цепь нуклеосом | В каждой нуклеосоме молекула ДНК делает по 2 оборота вокруг белкового октамера. | 10 нм | В 6,2 раза |
| II. Нуклеомерная нить (хроматиновая фибрилла, или нить) | Это цепочка нуклеомеров, содержащих по 5-8 нуклеосом, закрученных в суперспираль. | 30 нм | В 6-7 раз (всего - примерно в 40 раз) |
| III. Хромонема (хромосомная фибрилла) | Хроматиновая нить образует петли, собирающиеся в розетки, плотно прилегающие друг к другу. | 300 нм | Примерно в 250 раз (всего - в 10000 раз) |
| IV. Хроматида | Хромонема складывается в петли протяжённостью в 1 сегмент хроматиды. | 700 нм |
| Метафазная хромосома | Хромосома состоит из двух связанных друг с другом хроматид. | 1400 нм | Результи- рующее сокращение длины - как в хроматидах. |
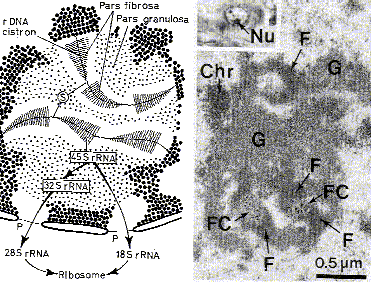
| Опреде- ление | Ядрышковые организаторы - это те участки хромосом, которые содержат гены рибосомных РНК. |
| Гены рРНК | а) Напомним: всего имеется 4 вида рРНК (п. 3.3.1.1). б) Гены трёх из них (28S, 18S и 5,8S) расположены рядом друг с другом, образуя кластер, транскрибируемый как единое целое. в) Ген четвёртой, самой короткой, РНК (5S) располагается и транскрибируется отдельно. |
| Количе- ство и локализа- ция генов рРНК | а) Указанный кластер, а также ген 5S-рРНК, во-первых, повторяются определённое число раз (образуя тандемы), и, во-вторых, содержатся (в виде тандемов) сразу в пяти парах хромосом: 13, 14, 15, 21 и 22 - а именно в области вторичной перетяжки этих хромосом. б) Таким образом, всего имеется 10 ядрышковых организаторов. в) Вместе они содержат у человека, по одним оценкам, около 200 копий каждого гена рРНК, а по другим – на порядок больше. |
| Количе- ство ядрышек | а) Количество ядрышек в ядре зависит от того, во сколько групп объединяются 10 ядрышковых организаторов – в одну, в две или больше. б) Таким образом, ядрышко – это не самостоятельное образование, а производное определённых хромосом. |
| Функция | Через поры в ядерной оболочке происходит интенсивный транспорт молекул и крупных частиц: а) в ядро из цитоплазмы – нуклеотидов (предшественников ДНК и РНК), хромосомных и рибосомных белков, регуляторных факторов и т.д.; б) из ядра в цитоплазму – многочисленных мРНК, тРНК, рибосомных субъединиц и пр. |
| Коли- чество пор | а) Поэтому количество пор в ядерной оболочке тем больше, чем интенсивней идут в клетке синтетические процессы. б) Обычно всего в оболочке ядра – 2000 – 4000 пор. |
| Отверстие поры | а) В области краёв поры (4) внутренняя и наружная мембраны сливаются, образуя округлое отверстие диаметром 120 нм. б) В отверстие встроен т.н. комплекс поры. | 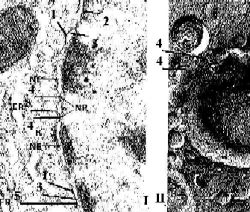 |
| Комплекс поры | а) Комплекс поры включает: тонкую диафрагму, закрывающую отверстие и пронизанную цилиндрическими каналами диаметром 10 нм; а также связанные с мембраной белковые гранулы. б) Из этих гранул одна расположена в центре комплекса, а 8 пар других – по периферии комплекса (с обеих сторон от диафрагмы). в) Центральная гранула связана фибриллами с периферическими. г) В итоге структура напоминает велосипедное колесо. |
| Переме- щение веществ через пору | а) Относительно небольшие молекулы диффундируют пассивно через гидрофильные каналы поры. б) А более крупные молекулы и частицы переносятся путём активного транспорта с затратой энергии ГТФ. Причём, в этом участвуют рецепторные белки, узнающие объект транспорта, и специальные белки, обеспечивающие (неизвестным пока способом) перемещение транспортного комплекса через диафрагму поры. в) По-видимому, центральная гранула – это и есть проходящий через пору транспортный комплекс. |
| Специаль- ный метод исследо- вания | а) Чтобы наблюдать внутреннюю поверхность ядерных мембран(II), используют особый способ приготовления препарата: замораживание и последующее скалывание, | |
| а затем - травление и напыление образца. б) В итоге, ядерные поры проявляются как округлые углубления. |
 2015-05-18
2015-05-18 2779
2779 4.1.1.2.Характеристика хромосомной ДНК I. Строение
4.1.1.2.Характеристика хромосомной ДНК I. Строение 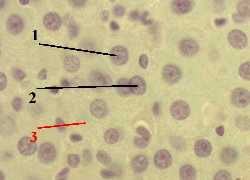 Полный размер
Полный размер  4.1.1.4. Выявление транскрипции в клеточных ядрах Все вышеназванные процессы могут быть выявлены с помощью специальных методик. Продемонстрируем это на примере транскрипции ДНК, т.е. матричного синтеза РНК. I. Принцип метода
4.1.1.4. Выявление транскрипции в клеточных ядрах Все вышеназванные процессы могут быть выявлены с помощью специальных методик. Продемонстрируем это на примере транскрипции ДНК, т.е. матричного синтеза РНК. I. Принцип метода  4.1.1.5. Структурные компоненты интерфазного ядра
4.1.1.5. Структурные компоненты интерфазного ядра 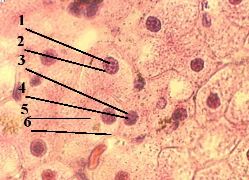 Полный размер
Полный размер  Полный размер
Полный размер 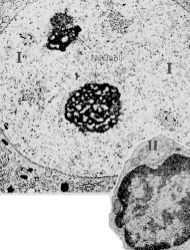 Полный размер
Полный размер 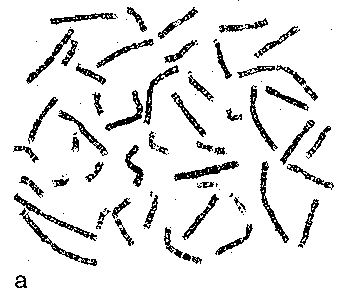
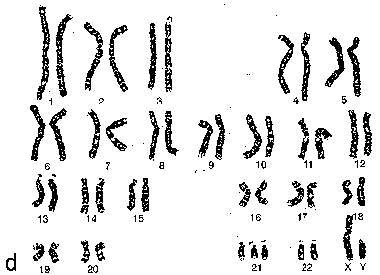
 Полный размер
Полный размер