 2015-05-10
2015-05-10 6949
6949Классическим методом идентификации интересующих участков ДНК стал метод блот-гибридизации по Саузерну, предложенный в 1975 г. Суть метода заключается в том, что сплошная "лестница" фрагментов ДНК, получившаяся в результате их деления по молекулярной массе в гелях, подвергается денатурации и переносится с геля на плотный носитель (нитроцеллю-лозный фильтр или нейлоновую мембрану). Перенос, или блоттинг, осуществляется за счёт капиллярных сил, электрического поля или вакуума (рис. 4-63). Фиксированную на фильтре ДНК гибридизуют с ДНК- или РНК-зондом, содержащим метку. Методом радиоавтографии определяют положение искомого фрагмента геномной ДНК на электрофореграмме. Блот-гибридизация
- высокочувствительный метод идентификации специфических последовательностей.
К настоящему времени разработано много модификаций этого метода. Так, ДНК-зонд не всегда метят радиоактивными изотопами, нередко используют соединения, ковалентно связывающиеся с ДНК; их можно обнаружить по образованию окрашенного продукта или флюоресценции. Длина олигонуклеотидов в ДНК-зондах также может сильно варьировать, будучи иногда очень короткой - в 15-20 пар нуклеотидов. Описаны методы дот- (пятно) или слот- (полоска) гибридизации, когда на твёрдый носитель наносят препараты ДНК или РНК без предварительной рестрикции или электрофореза и гибридизуют их с мечеными ДНК-зондами.
В. Установление первичной структуры ДНК-фрагментов (секвенирование ДНК)
Наиболее часто для установления первичной структуры ДНК используют дидезоксисеквени-рование. В реакционных пробах, содержащих денатурированную однонитевую ДНК, ДНК-полимеразу, дезоксинуклеозидтрифосфаты (дНТФ) - дАТФ, дЦТФ, дГТФ и дТТФ, один из которых является радиоактивным, и праймер инициируют синтез ДНК в присутствии специфических дидезоксинуклеозидтрифосфатов (ддНТФ), или терминаторов, - ддАТФ, ддЦТФ, ддГТФ или ддТТФ. Синтез одновременно ведут в четырёх параллельных пробах, в каждую из которых наряду с компонентами реакционной смеси прибавляют один из 4 ддНТФ. ддНТФ будут конкурировать с нормальными дНТФ за включение в растущую полинуклеотидную цепь. При встраивании ддНТФ вместо соответствующего нуклеотида синтез ДНК прекращается. В результате в каждой из пробирок получается набор различающихся по длине меченых фрагментов ДНК с одним из специфических дидезоксинуклеотидов на конце. После одновременного разделения этих фрагментов в электрическом поле на 4 соседних дорожках и радиоавтографии размер синтезированных молекул может быть установлен, а это значит, что может быть определена локализация терминирующих дидезоксинуклеотидов. На основании этих данных устанавливают последовательность нуклеотидов во вновь сингезированных фрагментах, комплементарных ДНК-матрице (рис. 4-64).
В настоящее время создают приборы для автоматического одновременного секвенирования большого числа проб с использованием меченных разными флюорохромами дидезоксинуклеотидов.
В то же время разрабатывают новые, более эффективные и экономичные методы секвенирования. Сущность одного из них заключается в следующем: из 4 нуклеозидтрифосфатов (дАТФ, дГТФ, дТТФ, дЦТФ) создаётся набор олигонуклеотидов (например, октануклеотидов), включающий все возможные варианты последовательностей. Эти октануклеотиды иммобилизуют (пришивают) в ячейках, в результате чего создаётся так называемая олигонуклеотидная
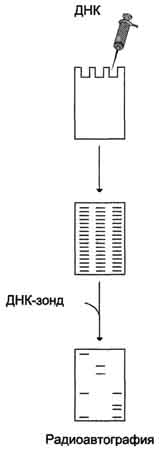
Рис. 4-63. Блот-гибридизация по Саузерну. Фрагменты ДНК разделяют с помощью электрофореза, денатурируют, переносят на нитроцеллюлозный фильтр и гибридизуют с ДНК-зондом.
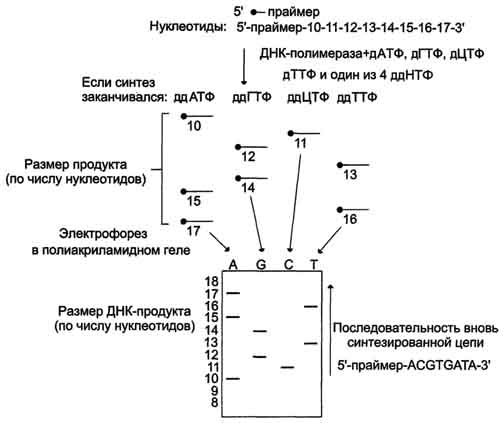
Рис. 4-64. Дидезоксисеквенирование последовательности ДНК. Используют 4 пробы, каждая из которых содержит ДНК-матрицу, праймер, ДНК-полимеразу, 4 дНТФ (дАТФ, дГТФ, дЦТФ, дТТФ). Праймер либо один из нуклеотидов содержит радиоактивную метку, благодаря чему полосы могут быть обнаружены в геле с помощью радиоавтографии. Один из 4 дидезоксирибонуклеотидов (ддНТФ) добавляют в каждую пробирку. Остановка синтеза происходит в том случае, когда ддНТФ включается в растущую олигонуклеотидную цепь.
матрица. Секвенируемый фрагмент ДНК метят по фосфатному остатку и добавляют в ячейки матрицы. Фрагмент ДНК гибридизуется только с теми октануклеотидами, последовательности которых комплементарны его участкам. Таким образом, определяется набор всех возможных октануклеотидов, присутствующих в исследуемом фрагменте ДНК. Далее при помощи специальной компьютерной обработки упорядочивается расположение октамеров в исследуемом фрагменте ДНК.
Г. Получение рекомбинантных ДНК и их амплификация
Для работы с нуклеотидными последовательностями в генах и других участках ДНК необходимо иметь достаточное количество материала для исследования. Это непростая задача, особенно если источником ДНК служат ткани человека. Поэтому исследуемые фрагменты ДНК обычно предварительно амплифицируют (увеличивают количественно в миллионы раз), для того чтобы получать их в любое время и в неограниченном количестве. Исключительно ценным инструментом в решении этой проблемы оказалось использование рекомбинантньгх ДНК (т.е. ДНК, построенных из участков разного происхождения).
1. Получение рекомбинантных ДНК
Для получения таких молекул первоначально выделяют ДНК из 2 разных источников (рис. 4-65).
Каждую из них в отдельности фрагментируют, используя одну и ту же рестриктазу, расщепляющую ДНК с образованием "липких" концов. После процедуры нагревания и медленного охлаждения (отжига) наряду с исходными
молекулами ДНКX и ДНКY могут образовываться рекомбинантные молекулы, состоящие из фрагментов ДНКX и ДНКY связанных между собой "липкими" концами. Ковалентное сшивание фрагментов осуществляют с помощью ДНК-лигазы в присутствии АТФ как источника энергии.
В технологии рекомбинантных ДНК, кроме фрагментов ДНК, выделенных из клеток, содержащих ядра, используют ДНК, полученную с помощью обратной транскриптазы. При добавлении в реакционную среду 4 разных дезоксирибонуклеозидтрифосфатов фермент на матрице мРНК по принципу комплементарности синтезирует ДНК-копию, или кДНК. Так как источником информации при образовании кДНК служит зрелая цитоплазматическая мРНК, то такая ДНК, в отличие от ДНК фрагментов, полученных при расщеплении геномной ДНК эукариотов, не содержит нитронов.








