 2015-05-10
2015-05-10 1189
1189Популяция ¾ совокупность особей одного вида, существующих в одно и то же время и занимающих определенную территорию.
Взаимодействие особей внутри популяции определяется внутривидовой конкуренцией, взаимодействие между популяциями ¾ межвидовой конкуренцией.
Внутривидовая конкуренция в популяции с дискретным размножением. Для популяций с дискретным размножением (некоторые виды растений, насекомых и т.д.) поколения четко разнесены во времени и особи разных поколений не сосуществуют. Численность такой популяции можно характеризовать числом N t и считать t величиной дискретной ¾ номером популяции.
Одна из моделей межвидовой конкуренции в этом случае выражается уравнением
 (1)
(1)
Здесь R ¾ скорость воспроизводства популяции в отсутствии внутривидовой конкуренции (математически это соответствует случаю a = 0). Тогда уравнение определяет просто изменение численности популяции по закону геометрической прогрессии:  , где N 0 ¾ начальная численность популяции.
, где N 0 ¾ начальная численность популяции.
Знаменатель в уравнении отражает наличие конкуренции, делающей скорость роста тем меньше, чем больше численность популяции; a и b ¾ параметры модели.
Исходные параметры модели:
· R ¾ скорость воспроизводства;
· N 0 ¾ начальная численность популяции;
· a ¾ параметр, характеризующий интенсивность внутривидовой конкуренции.
Характерная черта эволюции при b =1 ¾ выход численности популяции на стационарное значение при любых значениях других параметров. Однако, в природе так бывает не всегда, и более общая модель при b¹ 1 отражает другие, более сложные, но реально существующие, виды эволюции. Этих видов модель описывает четыре:
1) монотонное установление стационарной численности популяции;
2) колебательное установление стационарной численности популяции;
3) устойчивые предельные циклы изменения численности популяции;
4) случайные изменения численности популяции без наличия явных закономерностей (динамический хаос).
Внутривидовая конкуренция в популяции с непрерывным размножением. Математическая модель в данном случае строится на основе дифференциальных уравнений. Наиболее известна так называемая логистическая модель:
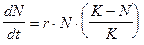 (2)
(2)
Исходные параметры модели:
· r ¾ скорость роста численности популяции в отсутствие конкуренции;
· K ¾ предельное значение численности популяции, при котором скорость роста становится равной нулю;
· N 0 ¾ начальная численность популяции.
Межвидовая конкуренция. В этом случае исследуется конкуренция популяций, потребляющих общий ресурс. Пусть N 1 и N 2 ¾ численности конкурирующих популяций. Модель (называемая также моделью Лотки-Вольтерры) выражается уравнениями
 (3)
(3)
Содержательный смысл параметров можно понять из сравнения с предыдущей моделью. Дополнительные параметры a 12и a 21 отражают интенсивность межвидовой конкуренции.
Главный вопрос, который интересует исследователя межвидовой конкуренции ¾ при каких условиях увеличивается или уменьшается численность каждого вида? Данная модель предсказывает следующие режимы эволюции взаимодействующих популяций: устойчивое сосуществование или полное вытеснение одной из них.
Система «хищник-жертва». В этой системе ситуация значительно отличается от предыдущей. В частности, если в случае конкурирующих популяций исчезновение одной означает выигрыш для другой (дополнительные ресурсы), то исчезновение «жертвы» влечет за собой и исчезновение «хищника», для которого в простейшей модели «жертва» является единственным кормом.
Обозначим через С численность популяции хищника и через N ¾ популяции жертвы. Одна из известных моделей выражается следующими уравнениями:
 (4)
(4)
В первое уравнение заложен следующий смысл. В отсутствии хищников (т.е. при С =0) численность жертв растет экспоненциально со скоростью r, т.к. модель не учитывает внутривидовой конкуренции. Скорость роста числа жертв (т.е. 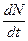 ) уменьшается тем больше, чем чаще происходят встречи представителей видов; а ¾ коэффициент эффективности поиска.
) уменьшается тем больше, чем чаще происходят встречи представителей видов; а ¾ коэффициент эффективности поиска.
Второе уравнение говорит о следующем. В отсутствии жертв численность хищников экспоненциально убывает со скоростью q; положительное слагаемое в правой части уравнения компенсирует эту убыль; f ¾ коэффициент эффективности перехода пищи в потомство хищников.








