 2015-05-26
2015-05-26 1344
1344Корневая система растений поглощает из почвы как воду, так и питательные минеральные вещества. Оба эти процесса взаимосвязаны, но осуществляются на основе разных механизмов. Корни извлекают минеральные вещества из почвенного раствора и из почвенного поглощающего комплекса, с частицами которого зона поглощения корня (корневые волески) тесно контактирует.
 Клеточные стенки принимают непосредственное участие как в поглощении веществ из почвы, так и в транспорте элементов минерального питания по тканям.
Клеточные стенки принимают непосредственное участие как в поглощении веществ из почвы, так и в транспорте элементов минерального питания по тканям.
Основной движущей силой поглотительной активности корней, как и вообще каждой клетки в отдельности, является работа ионных насосов (помп), локализованных в мембранах. Ра-
диальный транспорт минеральных веществ от поверхности корня к проводящей системе осуществляется в результате взаимодействия всех основных тканей зоны поглощения, причем каждая ткань выполняет определенные функции. Радиальный транспорт завершается загрузкой минеральных веществ и их органических производных в трахеиды и сосуды ксилемы. В другие части растения ксилемный сок перемещается за счет транспирации и (или) корневого давления (см. 5.4; 8.1). Клетки, входящие в состав различных тканей и органов, в свою очередь поглощают и метаболизируют элементы минерального питания, доставляемые с ксилемным соком. Причем их поглотительная активность зависит от возраста и функционального состояния.
В целом процесс минерального питания растения — это сложная цепь биофизических, биохимических и физиологических процессов со своими обратными и прямыми связями и системой регуляции. В настоящее время не все звенья этой цепи изучены достаточно подробно.
Поглотительная активность корня основывается на механизмах поглотительной активности, присущей любой растительной клетке. Поэтому такие общие вопросы, как избирательное поступление веществ в клетку, роль фазы клеточных стенок, трансмембранного переноса ионов будут обсуждаться применительно ко всем растительным клеткам.
Избирательное накопление минеральных веществ в клетке. В различных органах растений накапливается неодинаковое количество минеральных элементов, причем содержание минеральных веществ в клетках не соответствует концентрации этих же веществ во внешней среде. Например, данные табл. 6.1 показывают, что в тканях актинидии китайской концентрация фосфора в 1 — 10 тыс. раз превышает содержание этого элемента в почвенном растворе.
Таблица 6.1
| Сравнительное содержание элементов в тканях Actinidia chinensis (в ммоль/кг сырой массы плодов и листьев) и почвенном растворе (ммоль/л) (по R. L. Bieleski, I. В. Ferguson, 1983) | |||||
| Элементы | N | К | Р | Са | Mg |
| Ткани | |||||
| Почвенный раствор | 1-3 | 1-2 | 0,0005-0,002 | 0,5-1,5 | 2-4 |
В десятки раз выше в клетках содержание азота и калия. Это свидетельствует о том, что в клетках существуют механизмы не только поглощения веществ против концентрационного градиента, но и способы избирательного их накопления. Этот процесс начинается уже в клеточной стенке и затем продолжается с участием мембран.
Роль клеточных стенок в процессах адсорбции минеральных веществ. В отличие от клеток животных растительная клетка имеет оболочку (стенку), состоящую из целлюлозы, гемицеллюлоз и пектиновых веществ.
Пектиновые вещества (полиуро-новые кислоты) в своем составе содержат карбоксильные группы, в результате чего клеточные оболочки приобретают свойства катионообменников и могут концентрировать положительно заряженные вещества.
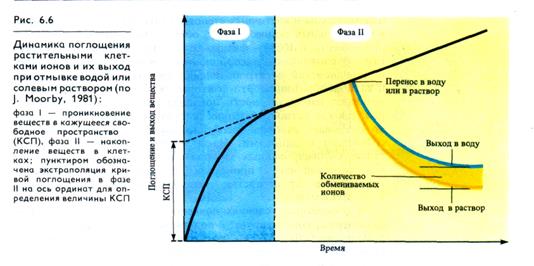
Если в сосуд, содержащий раствор 86RbCl или катионный краситель (например, метиленовый синий), погрузить корни (или другую растительную ткань), то в первые же 2 мин из раствора исчезнет до 50% рубидия (или красителя) от того количества, которое поглотится за длительное время (рис. 6. 6). В последующие 10 — 30 мин поглотится 70%, а дальнейшее связывание вещества тканями будет происходить очень медленно (часами). Чем обусловлено такое быстрое перемещение вещества в самом начале? Если ткань, находившуюся несколько часов в опытном растворе, перенести в воду или в солевой раствор того же состава, но без радиоактивной метки (или без красителя), то наблюдается обратная картина: быстрое выделение вещества в первые минуты и последующий медленный его выход из ткани. Таким образом, можно выделить две фазы поглощения веществ, протекающие с различными скоростями — высокой и медленной, причем вещество, быстро поглощенное тканью, так же быстро и выходит из нее. Первоначальное быстрое поглощение веществ осуществляется в клеточных стенках и является обменной адсорбцией (а быстрая потеря — десорбцией). Медленная фаза связана с функциональной активностью плазмалеммы (проникновением веществ в клетку или выходом из нее). Как отмечалось в разделе 1.12, молекулярное пространство в клеточной стенке, где происходят процессы обменной адсорбции, получило название кажущегося свободного пространства (КСП). Термин «кажущееся» означает, что объем этого свободного пространства зависит от объекта и природы растворенного вещества. КСП включает в себя межмолекулярное пространство в толще клеточных стенок и на поверхности плазмалеммы и клеточных стенок. По расчетам КСП занимает в растительных тканях 5—10% объема. Поглощение и выделение веществ в КСП — физико- химический пассивный процесс. Его обусловливают адсорбционные свойства ионообменника и доннановский электропотенциал на границе водной среды и катионообменника. Эти факторы уже на первом этапе обеспечивают избирательность поглощения веществ, несущих заряд, так как катионообменник (клеточные стенки) более активно связывает катионы (особенно двух- и трехвалентные) по сравнению с анионами. Из-за высокой плотности отрицательных фиксированных зарядов в клеточной стенке (1,4—1,8 мэкв/мг сухой массы) происходит первичное концентрирование катионов в пространстве, непосредственно примыкающем к плазмалемме.
В конкретных условиях почвенного питания клетки корня (ризодерма) контактируют с водной фазой (почвенным раствором) и с частицами почвы, которые также являются преимущественно катионообменниками (почвенный поглощающий комплекс). При этом большая часть минеральных питательных веществ находится не в растворе, а адсорбирована на частицах почвы.
Катионы и анионы поступают в клеточные стенки ризодермы как непосредственно из почвенного раствора, так и благодаря контактному обмену с частицами почвенного поглощающего комплекса. Оба эти процесса связаны с обменом ионов Н+ на катионы окружающей среды и HCO3(OH-) или анионов органических кислот на анионы минеральных веществ.
 Контактный обмен ионов клеточной стенки ризодермы (ионов Н+) с частицами почвы осуществляется без перехода ионов в почвенный раствор. Тесный контакт обеспечивается благодаря выделению слизи корневыми волосками и отсутствию у ризодермы кутикулы и других защитных покровных образований. Зона поглощения корней и частицы почвы образуют единую коллоидную систему (рис. 6. 7). Так как адсорбированные ионы находятся в постоянном колебательном движении и занимают определенный «осцилляторный объем» (сферу колебаний), при тесном контакте поверхностей сферы колебаний двух ближайших адсорбированных ионов могут перекрываться, в результате чего осуществляется ионный обмен.
Контактный обмен ионов клеточной стенки ризодермы (ионов Н+) с частицами почвы осуществляется без перехода ионов в почвенный раствор. Тесный контакт обеспечивается благодаря выделению слизи корневыми волосками и отсутствию у ризодермы кутикулы и других защитных покровных образований. Зона поглощения корней и частицы почвы образуют единую коллоидную систему (рис. 6. 7). Так как адсорбированные ионы находятся в постоянном колебательном движении и занимают определенный «осцилляторный объем» (сферу колебаний), при тесном контакте поверхностей сферы колебаний двух ближайших адсорбированных ионов могут перекрываться, в результате чего осуществляется ионный обмен.
Способность к обменной адсорбции вообще и контактному обмену в частности определяется обменной емкостью корня. Она зависит от химического состава корневых выделений и клеточных оболочек и поддерживается непрерывным синтезом новых веществ, связанным с ростом корня и с процессами обновления его структур, а также с поглощением веществ через цитоплазматическую мембрану внутрь клеток и дальнейшим их перемещением внутрь корня. Обменная емкость корня у разных видов растений неодинакова и зависит от возраста.

