 2015-05-26
2015-05-26 10903
10903«...все растения обладают способностью растворять белковые вещества... Это должно происходить при помощи растворителя, вероятно, состоящего из фермента вместе с кислотою».
Ч. Дарвин
Автотрофные организмы (от греч. «autos» — сам и «trophe» — питание) способны самостоятельно синтезировать органические питательные вещества из неорганических, гетеротрофные — питаются готовыми органическими веществами. К автотрофам принадлежат зеленые растения и некоторые бактерии, использующие в ходе фотосинтеза энергию света (фототрофы), а также бактерии, способные утилизировать энергию окисления веществ для синтеза органических соединений (хемосинтез).
Огромное большинство организмов, принадлежащих к царству растений, автотрофы (фототрофы). К гетеротрофам относят всех животных, грибы и большинство бактерий. Среди растений также имеются факультативные или облигатные гетеротрофы, получающие органическую пищу из внешней среды, — сапротрофы, паразиты и насекомоядные растения. Сапротрофы (сапрофиты) питаются органическими веществами разлагающихся остатков растений и животных, паразиты — органическими веществами живых организмов. Насекомоядные растения способны улавливать и переваривать мелких беспозвоночных. Однако в жизни растения есть периоды, когда оно питается только за счет запасенных ранее органических веществ, т. е. гетеротрофно. К таким периодам относятся прорастание семян, органов вегетативного размножения (клубней, луковиц и др.), рост побегов из корневищ, развитие почек и цветков у листопадных древесных растений и т. д. Многие органы растений гетеротрофны полностью или частично (корни, почки, цветки, плоды, формирующиеся семена). Наконец, все ткани и органы растения гетеротрофно питаются в темноте. Именно поэтому в культуре можно выращивать изолированные растительные клетки и ткани без света на органоминеральной среде.
Таким образом, гетеротрофный способ питания клеток и тканей столь же обычен для растений, как и фотосинтез, поскольку присущ любой клетке. В то же время этот способ питания растений изучен крайне недостаточно. Знакомство с физиологией растений, питающихся гетеротрофно, позволяет ближе подойти к пониманию механизмов питания клеток, тканей и органов в целом растении.
 Целые растения или органы могут усваивать как низкомолекулярные органические соединения, поступающие извне или из собственных запасных фондов, так и высокомолекулярные белки, полисахариды, а также жиры, которые необходимо предварительно перевести в легкодоступные и усвояемые соединения. Последнее достигается в результате пищеварения, под которым понимают процесс ферментативного расщепления ма-кромолекулярных органических соединений на продукты, лишенные видовой специфичности и пригодные для всасывания и усвоения. Различают три типа пищеварения: внутриклеточное, мембранное и внеклеточное. Внутриклеточное — самый древний тип пищеварения. У растений оно происходит не только в цитоплазме, но и в вакуолях, пластидах, белковых телах, сферосомах. Мембранное пищеварение осуществляется ферментами, локализованными в клеточных мембранах, что обеспечивает максимальное сопряжение пищеварительных и транспортных процессов. Оно хорошо изучено в кишечнике ряда животных. У растений мембранное пищеварение не исследовалось. Внеклеточное пищеварение происходит тогда, когда гидролитические ферменты, образующиеся в специальных клетках, выделяются в наружную среду и действуют вне клеток. Этот тип пищеварения характерен для насекомоядных растений; он осуществляется и в других случаях, в частности в эндосперме зерновок злаков.
Целые растения или органы могут усваивать как низкомолекулярные органические соединения, поступающие извне или из собственных запасных фондов, так и высокомолекулярные белки, полисахариды, а также жиры, которые необходимо предварительно перевести в легкодоступные и усвояемые соединения. Последнее достигается в результате пищеварения, под которым понимают процесс ферментативного расщепления ма-кромолекулярных органических соединений на продукты, лишенные видовой специфичности и пригодные для всасывания и усвоения. Различают три типа пищеварения: внутриклеточное, мембранное и внеклеточное. Внутриклеточное — самый древний тип пищеварения. У растений оно происходит не только в цитоплазме, но и в вакуолях, пластидах, белковых телах, сферосомах. Мембранное пищеварение осуществляется ферментами, локализованными в клеточных мембранах, что обеспечивает максимальное сопряжение пищеварительных и транспортных процессов. Оно хорошо изучено в кишечнике ряда животных. У растений мембранное пищеварение не исследовалось. Внеклеточное пищеварение происходит тогда, когда гидролитические ферменты, образующиеся в специальных клетках, выделяются в наружную среду и действуют вне клеток. Этот тип пищеварения характерен для насекомоядных растений; он осуществляется и в других случаях, в частности в эндосперме зерновок злаков.
В настоящее время грибы относят к самостоятельному царству, однако многие стороны физиологии грибов близки к физиологии растений. По-видимому, сходные механизмы лежат и в основе их гетеротрофного питания.
В плазмалемме гиф грибов, клеток дрожжей функционирует Н+-помпа, и в окружающую среду гриб выделяет различного рода кислые гидролазы. Это приводит к гидролизу сложных органических соединений в окружающем субстрате (внеклеточное кислое пищеварение) и всасыванию продуктов их распада.
Механизм всасывания также связан с работой Н+-помпы в плазмалемме. При закислении наружной примембранной зоны снижается диссоциация органических кислот и они проникают в клетки в виде нейтральных молекул. Сахара и аминокислоты поступают в цитоплазму гифы в симпорте с ионами Н+ с помощью специальных липопротеиновых переносчиков. Источником энергии для переноса органических веществ в симпорте с ионами Н+ служит протондвижущая сила, включающая как ΔрН, так и электрический мембранный потенциал, возникающие благодаря работе Н+-помпы в плазмалемме (см. рис. 6.8).
Среди растений сапрофитный способ питания довольно обычен у водорослей. Например, диатомовые водоросли, живущие на больших глубинах, куда не достигает свет, питаются, поглощая органические вещества из окружающей среды. При большом количестве растворимых органических веществ в водоемах легко переходят к гетеротрофному способу питания хлорококковые, эвгленовые и некоторые другие водоросли. Причем и в этом случае перенос Сахаров в клетки осуществляется в симпорте с ионами Н+, т. е. с использованием про-тондвижущей силы плазмалеммы.
У покрытосеменных растений сапрофитный способ питания относительно редок. Такие растения не имеют или имеют мало хлорофилла и не способны к фотосинтезу, хотя встречаются и фотосинтезирующие виды. Для построения своего тела они используют гниющие остатки растений и животных. Как пример можно привести Gidiophytum formicarum — полукустарник, стебель которого образует крупный клубень, пронизанный многочисленными ходами, в которых поселяются муравьи. Этот вид использует в пищу продукты жизнедеятельности муравьев, что было доказано с помощью радиоактивной метки. Меченые личинки мухи, которых муравьи занесли в полость стебля, были переварены растением через месяц, а радиоактивность была обнаружена в листьях и подземных частях растения.
 Некоторые виды, не содержащие хлорофилла, для обеспечения себя органической пищей используют симбиоз с грибами; это микотрофные растения. Особенно много таких видов в семействе орхидных. На ранних этапах развития все орхидеи вступают в симбиоз с грибами, так как запаса питательных веществ в их семенах недостаточно для роста зародыша. Гифы грибов, проникающие в семена, поставляют растущему зародышу органические вещества, а также минеральные соли из перегноя. У взрослых орхидей с микотрофным типом питания гифы грибов внедряются в периферическую зону корней, но дальше проникнуть не могут. Их дальнейшему росту препятствует фунгистатическое действие клеток глубинных тканей корня, а также слой довольно больших клеток с крупными ядрами, похожих на фагоциты. Эти клетки способны переваривать гифы грибов и усваивать освобождающиеся органические вещества. Возможен, вероятно, и прямой обмен между растением и грибом через наружную мембрану гифы.
Некоторые виды, не содержащие хлорофилла, для обеспечения себя органической пищей используют симбиоз с грибами; это микотрофные растения. Особенно много таких видов в семействе орхидных. На ранних этапах развития все орхидеи вступают в симбиоз с грибами, так как запаса питательных веществ в их семенах недостаточно для роста зародыша. Гифы грибов, проникающие в семена, поставляют растущему зародышу органические вещества, а также минеральные соли из перегноя. У взрослых орхидей с микотрофным типом питания гифы грибов внедряются в периферическую зону корней, но дальше проникнуть не могут. Их дальнейшему росту препятствует фунгистатическое действие клеток глубинных тканей корня, а также слой довольно больших клеток с крупными ядрами, похожих на фагоциты. Эти клетки способны переваривать гифы грибов и усваивать освобождающиеся органические вещества. Возможен, вероятно, и прямой обмен между растением и грибом через наружную мембрану гифы.
 По традиции такие бесхлорофилльные растения, как подъельник (Monotropa), также относят к сапрофитам. Однако и в этом случае сапрофитный способ питания осуществляется не непосредственно, а в симбиозе с грибами в форме микоризы. Причем во многих случаях эти симбиотические отношения можно рассматривать как форму паразитизма, когда клетки растений переваривают гифы гриба, проникшие в клетки корня. Таким образом, собственно сапротрофом является гриб, а высшее растение паразитирует на нем. Гифы гриба могут соединять корень подъельника с корнями дерева, и тогда подъельник становится паразитом, получающим органические вещества от другого растения.
По традиции такие бесхлорофилльные растения, как подъельник (Monotropa), также относят к сапрофитам. Однако и в этом случае сапрофитный способ питания осуществляется не непосредственно, а в симбиозе с грибами в форме микоризы. Причем во многих случаях эти симбиотические отношения можно рассматривать как форму паразитизма, когда клетки растений переваривают гифы гриба, проникшие в клетки корня. Таким образом, собственно сапротрофом является гриб, а высшее растение паразитирует на нем. Гифы гриба могут соединять корень подъельника с корнями дерева, и тогда подъельник становится паразитом, получающим органические вещества от другого растения.
Микориза большинством растений используется главным образом для увеличения поглощения воды и минеральных солей.
На примере подъельника и орхидей был рассмотрен способ питания высших растений путем паразитизма. Микоризный гриб также выступает как паразит (явление взаимного паразитизма). Гифы гриба образуют выросты-гаустории, плотно прилегающие к клеткам корней или же проникающие в них. Гаустории высасывают питательные вещества (прежде всего углеводы) из растения, и можно предполагать, что механизм этого явления также построен на базе Н+-помпы.
 Высшие растения-паразиты, использующие готовые органические вещества, — это, как правило, высокоспециализированные однолетники или многолетники с редуцированными или полностью утраченными в ходе эволюции листьями, а часто и корнями. Имеются виды, совершенно лишенные хлорофилла и не способные к фотосинтезу. К ним относится, например, заразиха (Orobanche), паразитирующая на корнях многих культурных растений. Ее семена прорастают лишь под влиянием корневых выделений растения-хозяина. Как только кончик зародышевого корня проростка соприкоснется с корнем хозяина, он преобразуется в гаусторию (присоску), начинающую выделять гидролазы, растворяющие клеточные стенки, и активно внедряющуюся в корень. В период роста и развития заразиха поглощает большое количество азотистых веществ, углеводов и минеральных элементов, особенно фосфора, а также воду из корней растений-хозяев. В растениях томатов, пораженных заразихой, содержание, например, белкового азота снижается в 3раза, а Сахаров — в 16 раз. Другой пример корневого паразита — петров крест (Lathraea squamaria), паразитирующий на корнях деревьев и кустарников.
Высшие растения-паразиты, использующие готовые органические вещества, — это, как правило, высокоспециализированные однолетники или многолетники с редуцированными или полностью утраченными в ходе эволюции листьями, а часто и корнями. Имеются виды, совершенно лишенные хлорофилла и не способные к фотосинтезу. К ним относится, например, заразиха (Orobanche), паразитирующая на корнях многих культурных растений. Ее семена прорастают лишь под влиянием корневых выделений растения-хозяина. Как только кончик зародышевого корня проростка соприкоснется с корнем хозяина, он преобразуется в гаусторию (присоску), начинающую выделять гидролазы, растворяющие клеточные стенки, и активно внедряющуюся в корень. В период роста и развития заразиха поглощает большое количество азотистых веществ, углеводов и минеральных элементов, особенно фосфора, а также воду из корней растений-хозяев. В растениях томатов, пораженных заразихой, содержание, например, белкового азота снижается в 3раза, а Сахаров — в 16 раз. Другой пример корневого паразита — петров крест (Lathraea squamaria), паразитирующий на корнях деревьев и кустарников.
У вьющегося паразитного травянистого растения повилики (Cuscuta) нитевидные стебли с редуцированными листьями-чешуйками обвиваются вокруг стеблей растений-хозяев и присасываются к ним с помощью гаусторий. Гаустории повилики — преобразованные адвентивные (придаточные) корни. Они принимают форму диска, плотно прилегающего к коре растения-хозяина. Группа клеток из центральной части диска внедряется в коровую паренхиму растения- хозяина и достигает центрального цилиндра, откуда повилика получает воду, органические вещества и минеральные элементы. Проростки повилики, совершая ростовые вращательные движения, находят растение-хозяина, реагируя на градиент влажности и выделяемые им вещества (явление хемотропизма — см. 13.5.2).
 К паразитным растениям относится и раффлезия, питающаяся соками корней тропических лиан. В тело жертвы она внедряется с помощью гаусторий, выделяющих целлюлазу и другие ферменты, разрушающие клеточные стенки. Всю свою жизнь раффлезия проводит в теле хозяина — под землей. Лишь ее цветки появляются на поверхности почвы. С помощью радиоактивной метки показано, что паразиты поглощают из тела хозяина в основном сахарозу, глутамимовую и аспарагиновую кислоты и их амиды.
К паразитным растениям относится и раффлезия, питающаяся соками корней тропических лиан. В тело жертвы она внедряется с помощью гаусторий, выделяющих целлюлазу и другие ферменты, разрушающие клеточные стенки. Всю свою жизнь раффлезия проводит в теле хозяина — под землей. Лишь ее цветки появляются на поверхности почвы. С помощью радиоактивной метки показано, что паразиты поглощают из тела хозяина в основном сахарозу, глутамимовую и аспарагиновую кислоты и их амиды.
В настоящее время известно свыше 400 видов покрытосеменных растений, которые ловят мелких насекомых и другие организмы, переваривают свою добычу и используют продукты ее разложения как дополнительный источник питания. Большинство из них встречается на бедных азотом болотистых почвах, есть эпифитные и водные формы.
 Ловчие механизмы. Листья насекомоядных растений трансформированы в специальные ловушки. Наряду с фотосинтезом они служат для поимки добычи. По способу ее ловли насекомоядные растения можно разделить на две большие группы.
Ловчие механизмы. Листья насекомоядных растений трансформированы в специальные ловушки. Наряду с фотосинтезом они служат для поимки добычи. По способу ее ловли насекомоядные растения можно разделить на две большие группы.
При пассивном типе ловли добыча может а) прилипать к листьям, желёзки которых выделяют липкую слизь, содержащую кислые полисахариды (библис, росолист), или б) попадать в специальные ловушки в виде кувшинов, урн, трубочек, окрашенных в яркие цвета и выделяющие сладкий ароматный секрет (саррацения, гелиамфора, дарлингтония).
Для активного захвата насекомых используются: 1) приклеивание добычи липкой слизью и обволакивание ее листом или волосками (жирянка, росянка), 2) ловля по принципу капкана — с захлопыванием ловчих листьев над добычей (аль-дрованда, венерина мухоловка), 3) ловчие пузырьки, в которые насекомые втягиваются с водой благодаря поддерживаемому в них вакууму (пузырчатка).
Общим для всех типов ловчих приспособлений является привлечение насекомых с помощью полисахаридных слизей или ароматного секрета (нектара), выделяемых или самими ловчими аппаратами, или желёзками вблизи от ловушки. Быстрые движения ловчих органов, как правило, осуществляются путем изменений в них тургора и запускаются с помощью распространяющихся потенциалов действия в ответ на раздражение чувствительных волосков, вызванное движениями насекомого (см. 13.6.1).
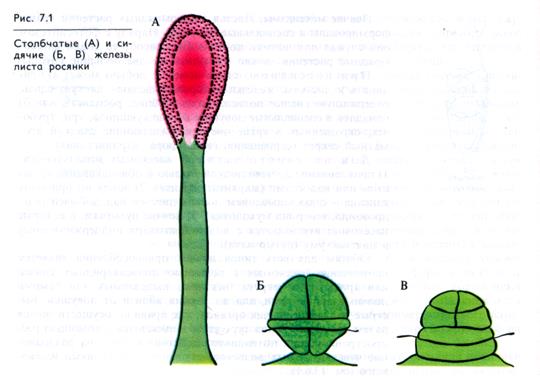
Пищеварение. Попавшее в ловушку или приклеившееся к липкой поверхности листа насекомое переваривается под действием секрета многочисленных желёзок (рис. 7.1). Некоторые насекомоядные парализуют добычу алкалоидами, содержащимися в выделяемой слизи. Так, росянка выделяет алкалоид ко-нитин, парализующий насекомое. Липкая слизь, секретируемая стоячими, или стебельчатыми, желёзками насекомоядных растений, содержит много кислых полисахаридов, состоящих из ксилозы, маннозы, галактозы и глюкуроновой кислоты. Кислая реакция слизи обеспечивается и находящимися в ней органическими кислотами. Слизь, выделяемая стоячими желёзками, содержит ряд гидролаз, активных в кислой среде. Например, у жирянки в слизи содержится кислая амилаза.
 Кислые слизистые выделения, азот- и фосфорсодержащие продукты распада стимулируют работу сидячих желёзок, которые начинают выделять кислоты (муравьиную, бензойную), а также протеазы и ряд других гидролаз. Довольно подробно изучена протеолитическая активность секрета у мухоловки. Большая часть протеаз из ее секрета относится к тиоловым протеазам, среди них — карбоксипептидаза, хитинолитическая активность. Кроме протеаз в секрете обнаружены кислая фос-фатаза и эстераза, а у некоторых видов насекомоядных — рибо-нуклеаза, липаза, а также пероксидаза.
Кислые слизистые выделения, азот- и фосфорсодержащие продукты распада стимулируют работу сидячих желёзок, которые начинают выделять кислоты (муравьиную, бензойную), а также протеазы и ряд других гидролаз. Довольно подробно изучена протеолитическая активность секрета у мухоловки. Большая часть протеаз из ее секрета относится к тиоловым протеазам, среди них — карбоксипептидаза, хитинолитическая активность. Кроме протеаз в секрете обнаружены кислая фос-фатаза и эстераза, а у некоторых видов насекомоядных — рибо-нуклеаза, липаза, а также пероксидаза.
Секреторные клетки насекомоядных растений имеют хорошо развитый ЭР и аппарат Гольджи, которые продуцируют большое количество секрета. Секреция слизи, кислот и гидролаз носит активный характер и зависит от дыхания. Дыхательные яды — арсенит, цианид, а также хлороформ прекращают процесс секреции и продуцирование пузырьков с секретом в аппарате Гольджи.
 Всасывание продуктов распада осуществляется теми же желёзками, соединенными с проводящей системой, и происходит довольно быстро, что было показано на примере мухоловки с помощью метиленовой сини. Через 5 мин краска проникала в цитоплазму всех клеток головки желёзок. Количественный ультраструктурный анализ показал доминирующую роль симпласта в транспорте продуктов пищеварения у мухоловки. Из всего сказанного следует, что процесс пищеварения у насекомоядных растений осуществляется принципиально так же, как в желудке животных. В том и другом случае секретируются кислоты (НС1 — в желудке, муравьиная кислота — у насекомоядных растений). Кислая реакция пищеварительного сока уже сама по себе способствует перевариванию животной пищи, так как в ее клетках активируются кислые лизосомные гидролазы. Выделение секреторными пищеварительными желёзками гидролаз (протеаз, фосфатаз, эстераз и др.), имеющих оптимум активности в кислой среде, создает благоприятные условия для переваривания пищи. На принципиальное сходство процесса кислого внеклеточного пищеварения у животных и растений впервые указал Дарвин в книге «Насекомоядные растения» (1875).
Всасывание продуктов распада осуществляется теми же желёзками, соединенными с проводящей системой, и происходит довольно быстро, что было показано на примере мухоловки с помощью метиленовой сини. Через 5 мин краска проникала в цитоплазму всех клеток головки желёзок. Количественный ультраструктурный анализ показал доминирующую роль симпласта в транспорте продуктов пищеварения у мухоловки. Из всего сказанного следует, что процесс пищеварения у насекомоядных растений осуществляется принципиально так же, как в желудке животных. В том и другом случае секретируются кислоты (НС1 — в желудке, муравьиная кислота — у насекомоядных растений). Кислая реакция пищеварительного сока уже сама по себе способствует перевариванию животной пищи, так как в ее клетках активируются кислые лизосомные гидролазы. Выделение секреторными пищеварительными желёзками гидролаз (протеаз, фосфатаз, эстераз и др.), имеющих оптимум активности в кислой среде, создает благоприятные условия для переваривания пищи. На принципиальное сходство процесса кислого внеклеточного пищеварения у животных и растений впервые указал Дарвин в книге «Насекомоядные растения» (1875).
Сходство физиологии пищеварения у животных и растений уже не вызывает сомнения. В настоящее время известно, что закисление среды в желудке животных осуществляется в результате функционирования Н+ помпы в плазмалемме клеток слизистой желудка. Вслед за ионами Н+ в полость желудка поступают анионы хлора, компенсирующие выход протонов. Для растений хлор не является необходимым макроэлементом и не накапливается в больших количествах в клетках. Поэтому вслед за ионами Н+ из секреторных клеток пищеварительных органов насекомоядных растений выходит не хлор, а анионы органических кислот, прежде всего муравьиная кислота. Свойства кислой протеазы насекомоядных растений также сходны с пепсином желудка, максимальная активность которого наблюдается при рН 1—2.
Значение насекомоядное™. Многие насекомоядные растения живут на почвах, бедных минеральными элементами. Их корневая система слабо развита. У этих растений в отличие от других болотных растений, как правило, нет микоризы, поэтому усвоение минеральных элементов из пойманной добычи имеет для них большое значение. Из тела жертвы насекомоядные растения получают азот, фосфор, калий, серу. Углерод, содержащийся в аминокислотах и других продуктах распада, также участвует в метаболизме насекомоядных растений. Еще Дарвин показал, что если растения росянки подкармливать кусочками мяса, то через три месяца они значительно превосходят контрольные растения по ряду показателей, особенно репродуктивных. Установлено, что растения пузырчатки зацветают только после получения животной пищи.
В основе свойства насекомоядных растений питаться плотоядно лежит способность любой растительной клетки использовать для своего питания органические вещества, освобождаемые из запасной формы или притекающие из других частей растения. В целом растительные организмы, как правило, автотрофны, но углеродные соединения, синтезируемые в ходе фотосинтеза из СО2и воды, затем поступают из листьев во все другие части растения, которые питаются за счет этих готовых органических веществ, т. е. гетеротрофно. В тех случаях, когда растительный организм использует запасные органические вещества (углеводы, белки, жиры) или биополимеры цитоплазмы (например, из стареющих листьев), эти вещества должны быть предварительно гидролизованы и таким образом превращены в транспортабельную и усвояемую форму. Этот процесс принципиально не отличается от пищеварения у насекомоядных растений.
Процессы пищеварения в эндосперме. Наиболее удобный объект для изучения процесса пищеварения у растений — прорастающие семена, имеющие эндосперм с отложенными в нем запасами органических питательных веществ. В зрелых зерновках злаков зародыш непосредственно не контактирует с тканями эндосперма. Для мобилизации и всасывания запасных органических веществ (полисахаридов, белков) служит видоизмененная семядоля — щиток (рис. 7.2). По существу, функциональная роль щитка аналогична функции желудка животных. Можно ожидать, что и механизм переваривания нерастворимых запасных полисахаридов и белков эндосперма при прорастании семян принципиально сходен с тем, что происходит при переваривании животных тканей у насекомоядных растений и с пищеварительными процессами в желудке животных.
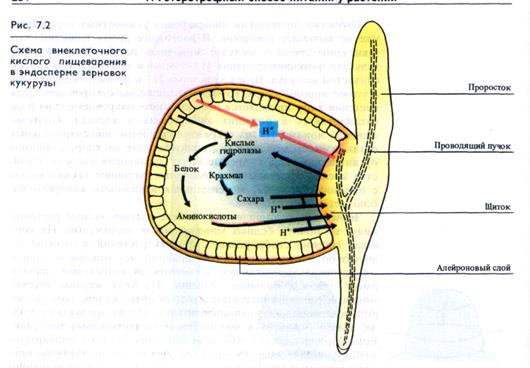
Действительно, в эпителиальных клетках щитка функционирует Н+-помпа, и щиток выделяет в эндосперм органические кислоты (лимонную и др.). В результате закисления клеток эндосперма, являющихся полумертвыми (ядра и многие органоиды в запасающих клетках эндосперма при созревании зерновок разрушаются), в них активируются кислые гидролазы, прежде всего α-амилазы, и крахмал разлагается до мальтозы и глюкозы. Вслед за секрецией кислот эпителиальные клетки щитка выделяют в эндосперм кислые гидролазы: α- и β-амилазы, цел-люлазу, протеазы, различные глюканазы, фосфатазу, РНКазу и др.
Вслед за щитком на третьи-четвертые сутки прорастания к пищеварительной деятельности щитка подключается и единственный живой слой клеток в эндосперме — периферический алейроновый слой (рис. 7.2). Алейроновые клетки также выделяют в эндосперм органические кислоты и кислые гидролазы (α-амилазу, протеазы, РНКазу и др.). Совместная деятельность щитка и алейронового слоя приводит к растворению запасных веществ эндосперма.
Щиток является также всасывающим органом. Плазмалеммой эпителиальных клеток щитка, граничащих с эндоспермом, осуществляется перенос Сахаров, аминокислот, неорганических катионов и анионов из эндосперма в цитоплазму клеток щитка с последующим поступлением транспортируемых питательных соединений в проводящие пучки, которые доставляют эти соединения в растущий зародыш. Этот процесс связан с работой мембранных Н+-помп, создающих протондвижущую силу. Перенос органических веществ через плазмалемму эпителиальных клеток щитка происходит в симпорте с ионами Н +.
Важно отметить, что процессы мобилизации запасных веществ в эндосперме находятся под гормональным контролем. Секреторная активность щитка активируется цитокинином и ауксином, а синтез и секреция гидролаз в алейроновых клетках полностью находятся под контролем гиббереллина, который поступает в алейроновые клетки из щитка и зародыша.
Растворение и отток запасных веществ из других запасающих органов, например из семядолей двудольных при прорастании семян, по-видимому, происходит сходным образом. Однако в этом случае осуществляется не внеклеточное, а внутриклеточное кислое пищеварение. Эти процессы изучены еще недостаточно.
Мобилизация запасного белка в прорастающих семенах. Сезонная периодичность развития растений включает периоды покоя, перед наступлением которых ассимиляты, оттекающие из листьев, откладываются в запас. Эти запасенные питательные вещества необходимы для начала роста в следующий вегетационный период. Запасающие ткани семян, корней, стеблей, клубней представляют собой вместилища резервов питательных веществ. Накопление в клетках большого количества азота, углерода и других элементов требует образования осмотически неактивных или слабоактивных компонентов. Таковыми и являются макромолекулярные формы запасных питательных веществ: белки, полисахариды, триглицериды. Содержание белка в семенах довольно высоко. Больше всего белка в семенах бобовых (20 — 30%) и масличных (17 — 42%). В семенах злаков его 7—14% на сухую массу. Запасные белки локализованы в алейроновых зернах и белковых телах и представлены глобулинами (основная форма у двудольных) и альбуминами. Имеются также проламины и глютелины.
 Алейроновые зерна — запасающие органеллы клеток размером от 0,1 до 25 мкм. Окружены одинарной мембраной. Содержание белка 70 — 80% на сухую массу. В состав алейроновых зерен входят также углеводы, фосфолипиды, фитин, РНК, соли щавелевой кислоты. Встречаются сложные и простые алейроновые зерна. Сложные алейроновые зерна характерны для некоторых двудольных и имеют включения двух типов — глобоидные и кристаллоидные, окруженные аморфным материалом. В кристаллоиде сосредоточено около 60%, в аморфной зоне — 35 —40 % и глобоиде — 3 — 5% белка от общего его содержания в алейроновых зернах. Часть запасных белков в алейроновых зернах образует комплексы с фитином, углеводами и липидами, которые при прорастании гидролизи-руются в первую очередь. В глобоиде локализован фитин — кальций- магниевая соль инозитфосфорной кислоты. В алейроновых клетках крахмалистого эндосперма злаков представлены простые алейроновые зерна. По размеру они меньше сложных и не содержат включений. Фитин локализован у них в белковом аморфном матриксе. В алейроновых зернах содержатся некоторые кислые гидролазы.
Алейроновые зерна — запасающие органеллы клеток размером от 0,1 до 25 мкм. Окружены одинарной мембраной. Содержание белка 70 — 80% на сухую массу. В состав алейроновых зерен входят также углеводы, фосфолипиды, фитин, РНК, соли щавелевой кислоты. Встречаются сложные и простые алейроновые зерна. Сложные алейроновые зерна характерны для некоторых двудольных и имеют включения двух типов — глобоидные и кристаллоидные, окруженные аморфным материалом. В кристаллоиде сосредоточено около 60%, в аморфной зоне — 35 —40 % и глобоиде — 3 — 5% белка от общего его содержания в алейроновых зернах. Часть запасных белков в алейроновых зернах образует комплексы с фитином, углеводами и липидами, которые при прорастании гидролизи-руются в первую очередь. В глобоиде локализован фитин — кальций- магниевая соль инозитфосфорной кислоты. В алейроновых клетках крахмалистого эндосперма злаков представлены простые алейроновые зерна. По размеру они меньше сложных и не содержат включений. Фитин локализован у них в белковом аморфном матриксе. В алейроновых зернах содержатся некоторые кислые гидролазы.
Белковые тела находятся в крахмалистом эндосперме зерновок злаков. Эти гетерогенные системы, включающие кроме различных групп запасных белков крахмал, липиды и ряд кислых гидролаз, имеют размеры в несколько микрометров.
Распад белков в семенах начинается почти сразу же после начала набухания и осуществляется несколькими группами протеаз. Различают три стадии протеолиза запасных белков при прорастании. На первой стадии идет лишь ограниченный протеолиз основной массы запасных белков. Распадаются альбумины и глобулины, локализованные в осевых органах зародыша, в алейроновых зернах алейронового слоя. Повышаются подвижность и растворимость белков. Гидролиз белков на этой стадии поставляет аминокислоты, необходимые для синтеза новых ферментативных белков. На второй стадии, которая длится 5—10 дней, белки в запасающих органах быстро распадаются до аминокислот, которые транспортируются к растущему зародышу, обеспечивая его гетеротрофное питание. На заключительном этапе в запасающих органах полностью деградируют структурные и ферментативные белки, которые обеспечивали процесс пищеварения.
Для семян двудольных характерно внутриклеточное переваривание белков. Их распад происходит в тех же клетках, в которых они отложены в запас. Значительные изменения в структуре алейроновых зерен заметны уже через 3 — 5 дней с начала набухания. В них резко возрастает активность кислых гидролаз — фосфатазы, протеазы. В первую очередь исчезают белки аморфного матрикса, затем белковые кристаллоиды и в последнюю очередь глобоиды. Каким образом поддерживается кислый уровень рН в алейроновых зернах, неизвестно. Возможно, в подкислении участвует обнаруженная в них щавелевая кислота. Для полного гидролиза белков в алейроновых зернах недостает собственных протеаз. Дополнительная эндопептида-за синтезируется в эндоплазматическом ретикулуме, который примыкает к алейроновым зернам. По мере истощения белковых запасов алейроновые зерна превращаются в вакуоли.
В зерновках злаков наряду с внутриклеточным пищеварением в алейроновом слое, щитке и зародыше осуществляется и внеклеточное пищеварение неживых клеток крахмалистого эндосперма. Внеклеточное пищеварение идет под действием щитка и клеток алейронового слоя, выделяющих в эндосперм  кислоты, а затем и ряд кислых гидролаз, в том числе протеазы (см. рис. 7.2). Выделение кислот в эндосперм приводит к его подкислению, чем создаются оптимальные условия для работы гидролаз, имеющих оптимум действия при рН 4 — 6, а также для транспортных систем в эпителиальных клетках щитка, прилегающих к эндосперму и осуществляющих транспорт продуктов гидролиза из эндосперма в проводящие сосуды щитка. Установлено, что аминокислоты и дипептиды транспортируются гораздо эффективнее при кислых значениях рН наружной среды. Секреция гидролаз щитками кукурузы также наиболее интенсивна при рН 5,4 — 6,0.
кислоты, а затем и ряд кислых гидролаз, в том числе протеазы (см. рис. 7.2). Выделение кислот в эндосперм приводит к его подкислению, чем создаются оптимальные условия для работы гидролаз, имеющих оптимум действия при рН 4 — 6, а также для транспортных систем в эпителиальных клетках щитка, прилегающих к эндосперму и осуществляющих транспорт продуктов гидролиза из эндосперма в проводящие сосуды щитка. Установлено, что аминокислоты и дипептиды транспортируются гораздо эффективнее при кислых значениях рН наружной среды. Секреция гидролаз щитками кукурузы также наиболее интенсивна при рН 5,4 — 6,0.
Таким образом, подкисление эндосперма — важный этап в прорастании зерновок, так как он настраивает всю систему кислого внеклеточного пищеварения и транспорта на рабочий ритм. Низкомолекулярные пептиды, образующиеся вместе с аминокислотами при гидролизе белков, поглощаются щитками и сразу же гидролизуются щелочной эндопептидазой до аминокислот.
В семядолях тыквенных и бобовых распад белков начинается в алейроновых зернах вблизи эпидермиса и проводящих пучков, затем в столбчатой паренхиме и в последнюю очередь — в клетках губчатой паренхимы. В эндосперме злаков белковые тела сначала распадаются вблизи щитка, потом в алейроновом слое. Заметная потеря азота в крахмалистом эндосперме наблюдается уже на вторые сутки с начала набухания. В клетках алейронового слоя гидролиз белка начинается позже, на 3 —5-е сутки.
Мобилизация запасенных углеводов. Углеводы — важнейшая группа запасных питательных веществ семян. Некоторые семена содержат небольшой запас сахарозы, а также другие сахара (в зависимости от вида): стахиозу, мальтозу, галактозу, рибо-зу, фруктозу, глюкозу. Часто эти сахара связаны с белками в гликопротеиновые комплексы. Однако основным запасным полисахаридом в семенах является крахмал. Семена злаков содержат 50 — 76% крахмала от сухой массы (кукуруза и сорго — до 76, пшеница и ячмень-до 70%), бобовые-50-60% (горох-до 50, бобы — до 60%). Крахмал откладывается в пластидах при созревании семян. Когда крахмальные зерна вырастают до полных размеров, ламеллярная структура пластид разрушается. Размеры крахмальных зерен в семенах различных растений варьируют в пределах примерно от 15 (рис) до 50 мкм в диаметре (бобы). Кроме Сахаров и крахмала в качестве резервных углеводов могут выступать полисахариды, входящие в состав клеточных стенок. Это прежде всего гемицеллюлозы (галакто-маннаны, глюкофруктаны и др.).
При прорастании быстро исчезают запасы свободных Сахаров. Крахмальные гранулы, находящиеся в тесном контакте с цитоплазмой, начинают распадаться. На их поверхности обнаруживается шероховатость, указывающая на начавшийся гидролиз. На ранних стадиях прорастания часть крахмала распадается под действием фосфорилазы. Постепенно скорость фосфоролиза уменьшается и включается другой, более мощный механизм расщепления крахмала с помощью амилаз — гидролитический. Чтобы полностью расщепить крахмал до глюкозы гидролитическим путем, требуется целый комплекс ферментов: важнейшие из них — α- и (β-амилазы, а также α-глюкозидаза и предельная декстриназа.
 Переваривание крахмала происходит как внутри клетки, так и внеклеточным путем. Внутриклеточное пищеварение характерно для семян двудольных растений и начинается уже в первые часы набухания в осевых органах. Содержание крахмала в семядолях гороха начинает снижаться лишь с 4 —5-го дня прорастания.
Переваривание крахмала происходит как внутри клетки, так и внеклеточным путем. Внутриклеточное пищеварение характерно для семян двудольных растений и начинается уже в первые часы набухания в осевых органах. Содержание крахмала в семядолях гороха начинает снижаться лишь с 4 —5-го дня прорастания.
Лучше всего ход распада крахмала изучен у зерновок хлебных злаков, где основная его масса вовлекается в обмен через внеклеточное пищеварение. Процесс гидролиза начинается от щитка и распространяется к дистальной части эндосперма. В зоне эндосперма, примыкающей к щитку, уже через 20 ч от начала набухания зерновок крахмальные зерна разрушаются. Эпителиальные клетки щитка выделяют α- и β-амилазы, сначала запасные формы, а затем и вновь синтезированные. Щиток обеспечивает до 17% всей амилазной активности эндосперма. Выделение з-амилазы клетками алейронового слоя в эндосперм начинается на 3 —4-е сутки. За этот срок крахмал эндосперма уже частично переварен, наблюдается деструкция клеточных стенок, что облегчает процесс секреции и пищеварение.
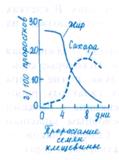
Усвоение жиров. Свыше 75% семян всех видов цветковых растений содержат жиры в качестве питательного резерва, используемого при прорастании. Запасы как нейтральных жиров, так и значительные количества полярных липидов сосредоточены в сферосомах. Это частицы сферической формы диаметром до 0,5 мкм, а иногда и больше, окруженные одинарной мембраной. В сферосомах обнаружена активность кислой липазы. Основные функции сферосом — запасание и переваривание жиров. У злаков этот процесс начинается сразу после начала набухания зерновок, хотя скорость его первоначально невысока и достигает максимума на 3 —4-й день от начала набухания. У семян масличных распад запасных жиров начинается на 2 —3-й сутки от начала набухания и проходит, как и у других групп растений, в три этапа. На первом этапе жиры под действием фермента липазы распадаются на глицерин и жирные кислоты; на втором — жирные кислоты расщепляются в ходе окисления до ацетил-СоА, который на третьем этапе может превращаться в другие соединения (см. 4.24 и 4.42) или подвергаться дальнейшему окислению.
Ключевая роль в переваривании жиров принадлежит кислой липазе, обладающей невысокой специфичностью и расщепляющей все глицериды. Она может депонироваться в созревающих семенах, а может синтезироваться вновь при прорастании семян в эндоплазматическом ретикулуме, а затем транспортироваться в сферосомы клетки. При прорастании семян отмечен тесный контакт сферосом с глиоксисомами, в которых под действием щелочной липазы происходит дальнейший распад моноглицеридов, а также β-окисление жирных кислот. Там же протекают реакции глиоксилатного цикла, в которых ацетил-СоА может превращаться в янтарную и щавелевоуксусную кислоты. Еще один продукт распада жиров — глицерин — восстанавливается до фосфодиоксиацетона и далее может превращаться в сахара путем глюконеогенеза (см. рис. 4.11).
Гетеротрофно могут питаться растения-сапрофиты, паразиты и насекомоядные растения. Пищеварительные желёзки насекомоядных растений выделяют кислоту (муравьиную), кислые гидролазы (про-теазы и др.) и осуществляют внеклеточное кислое пищеварение. Почти так же происходит мобилизация запасных веществ в эндосперме. Из щитка и алейроновых клеток зерновок злаков в эндосперм поступают органические кислоты и кислые гидролазы, а затем переваренные продукты всасываются щитком.
Важность изучения гетеротрофного питания определяется тем, что все незеленые органы растений (корни, цветки, плоды) и все клетки растений в темноте питаются гетеротрофно.



