 2015-06-16
2015-06-16 3195
3195Причины S–образной формы кривой диссоциации оксигемоглобина до конца не ясны. Если бы каждая молекула гемоглобина присоединяла только одну молекулуO2, то
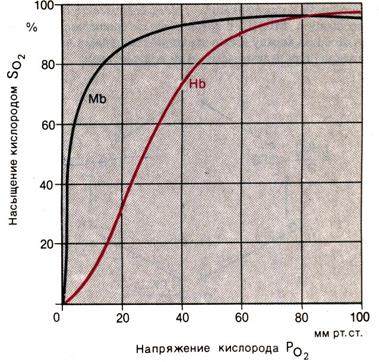
Рис. Кривые диссоциации оксигемоглобина (Hb) и оксимиоглобина (Mb) при рН 7,4 и t 37 °С
кинетика этой реакции графически описывалась бы гиперболой. Именно такая гиперболическая кривая диссоциации характерна, например, для реакции соединения кислорода с миоглобином (Mb), аналогичной реакции оксигенации гемоглобина. Строение миоглобина сходно со структурой одной из четырех субъединиц гемоглобина, поэтому молекулярные массы этих двух веществ соотносятся как 1:4. Поскольку в состав миоглобина входит лишь одна пигментная группа, одна молекула миоглобина может присоединить только одну молекулу O2:
Mb + O2 ↔ МbO2. (6)
Гиперболическая кривая диссоциации для этой реакции приведена на рис. 22.6. Исходя из вполне правдоподобного предположения о том, что S–образная форма кривой диссоциации НbO2 обусловлена связыванием одной молекулой гемоглобина четырех молекул O2, Эдер выдвинул так называемую гипотезу промежуточных соединений. Согласно этой гипотезе, присоединение четырех молекул O2 к гемоглобину происходит в несколько стадий, причем каждая из этих стадий влияет на равновесие следующей реакции. Таким образом, реакция соединения кислорода с гемоглобином описывается четырьмя константами равновесия, что и объясняет сигмоидную форму кривой диссоциации оксигемоглобина.
В то же время возможно и другое объяснение, согласно которому существуют две формы гемоглобина–оксигенированная и дезоксигенированная, переходящие одна в другую в результате конформационных перестроек. Если предположить, что параметры равновесия реакций оксигенации для этих двух форм гемоглобина различны, то с позиции данной гипотезы можно объяснить S–образную форму кривой диссоциации НbO2.
Биологический смысл формы кривой диссоциации оксигемоглобина. Конфигурация кривой диссоциации оксигемоглобина имеет важное значение с точки зрения переноса кислорода кровью. В процессе поглощения кислорода в легких напряжение O2 в крови (PO2) приближается к таковому в альвеолах. У молодых людей РO2 артериальной крови составляет около 95 мм рт.,ст. (12,6 кПа). Из рис. 22.6 видно, что при таком напряжении насыщение гемоглобина кислородом составляет примерно 97%. С возрастом (и в еще большей степени при заболеваниях легких) напряжение O2 в артериальной крови может значительно снижаться, однако, поскольку кривая диссоциации оксигемоглобина в правой ее части почти горизонтальна, насыщение крови кислородом уменьшается ненамного. Так, даже при падении РO2 в артериальной крови до 60 мм рт. ст. (8,0 кПа) насыщение гемоглобина кислородом равно 90%. Таким образом, благодаря тому что области высоких напряжений кислорода соответствует горизонтальный участок кривой диссоциации оксигемоглобина, предупреждается существенное снижение насыщения артериальной крови кислородом.
Крутой наклон среднего участка кривой диссоциации оксигемоглобина свидетельствует об очень благоприятных условиях для отдачи кислорода тканям. При изменении локальной потребности в кислороде он должен высвобождаться в достаточном количестве в отсутствие значительных сдвигов PO2 в артериальной крови. В состоянии покоя PO2 в области венозного конца капилляра равно приблизительно 40 мм рт. ст. (5,3 кПа), что соответствует примерно 73% насыщения. Если в результате увеличения потребления кислорода его напряжение в венозной крови падает лишь на 5 мм рт. ст. (0,7 кПа), то насыщение гемоглобина кислородом снижается не менее чем на 7%; высвобождающийся при этом O2 может быть сразу же использован для процессов метаболизма.
Содержание O2 в артериальной и венозной крови.
Количество химически связанного кислорода в крови зависит от насыщения им гемоглобина (SO2). Зная величину SO2, можно, исходя из числа Хюфнера, вычислить объемное содержаниеO2 в крови (в лO2 на 1л крови):
[O2]= 1,34·[Hb]·SO2·10–5 (7)
где SO2 выражено в процентах, a [Hb] – в граммах на литр. Подставляя в это уравнение количественные значения кислородного насыщения, можно вычислить, что в артериальной крови (SO2 = 97%) содержание кислорода составляет около 0,20, а в венозной (SO2 = 73%) оно равно 0,15. Таким образом, артериовенозная разница по концентрации кислорода (авРO2) составляет 0,05 (табл. 22.2). Это означает, что в норме при прохождении крови через тканевые капилляры используется лишь 25% общей кислородной емкости. Разумеется, разные органы существенно различаются по степени извлечения кислорода (см. рис. 23.2), так что величины для венозной крови, приведенные в табл., представляют собой средние значения показателей, варьирующих в широких пределах. При интенсивной физической нагрузке артериовенозная разница по кислороду может превышать 0,1.








