 2015-06-28
2015-06-28 1036
1036Эксперименты по составлению карт показали, что область rII состоит из многих мелких участков, или сайтов, в которых могут происходить разные мутации. Но такие карты дают представление только о строении гена и ничего не говорят о его функции. Даже неизвестно, состоит ли область rII из одного гена или нескольких. Для определения границ гена необходимы другие опыты, не имеющие ничего общего с кроссинговером и составлением карт даже если внешне эти опыты выглядят как эксперименты по составлению карт. Такие опыты называются комплементационными тестами, и их лучше всего объяснить при помощи модели.
Предположим, что мутации rII затрагивают два различных гена, которые расположены рядом, и что при мутации они оба дают одинаковый фенотип. Так как предполагается, что отдельный ген кодирует информацию о синтезе отдельного полипептида, то эти два гена должны кодировать синтез двух отдельных полипептидов, которые мы назовем А и В. Предположим, что оба гена необходимы для нормального функционирования в клетках К (для штамма В они несущественны). Тогда, если смешать клетки К с двумя различными мутантами, можно узнать, производятся ли оба белка. На рис. 8.2 показано, как различные мутации могут воздействовать на эти гены. Допустим, обе мутации происходят в гене А. Так как функциональный белок А не производится, то фаг расти не может. Теперь предположим, что одна мутация затрагивает ген А, а другая — ген В. Теперь в одном фаге имеется функциональный ген В, а в другом — функциональный ген А. Если клетку одновременно заразить этими двумя фагами, то они могут дополнить друг друга (то есть быть комплементарными друг другу): каждый выполняет функцию, отсутствующую у другого, и оба они могут расти. (Еще раз заметим, что эти тесты проверяют только функции генов, они не учитывают кроссинговер и рекомбинации.)
Когда Бензер заразил бактерии Е. coli К смесью мутантов rII, он получил именно те результаты, которые и предсказывала модель. Мутационные участки расположены вдоль линии и разделены на две группы.
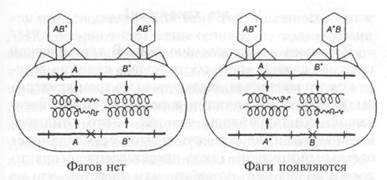
Рис. 8.2. С помощью комплементационного теста можно определить, происходят ли две мутации внутри одного гена или нет. Бактерии одновременно заражают двумя фагами с двумя различными мутациями, которые затрагивают либо один ген (слева), либо два гена (справа). Если мутации затрагивают один ген, то ни в одном фаге не создается нормальной копии гена, поэтому фаги не могут размножаться. Но если мутации затрагивают оба гена, то один фаг имеет нормальный ген А, а другой — нормальный ген В, и оба гена дополняют друг друга. Обратите внимание, что этот тест не имеет ничего общего с кроссинговером
Ни один из мутантов по левой группе не дополнял мутантов по этой же группе, и то же самое было с правой частью. В то же время любой мутант из левой группы оказывался комплементарным к любому мутанту из правой группы. Эти результаты доказывают, что область rII действительно включает в себя два гена. (Хотя Бензер называл отдельную функциональную единицу цистроном, сейчас цистроном называют то же, что и ген.) Комплементационные тесты, подобные этому, в наши дни применяют ко всем организмам, чтобы узнать, происходят ли две мутации внутри одного гена или нет, и определить таким образом границу между генами.
Что же такое ген!
Вернемся к определению гена. В классической генетике словом «ген» обозначалась единица генетического материала, выделяемая по трем критериям: по функции, мутации и рекомбинации. Изначально предполагалось, что ген — это функциональная единица, то есть нечто, определяющее отдельный признак. Такое представление сохранилось и до сих пор, но сейчас нам известно, что на один и тот же признак могут воздействовать различные гены и что при мутации гены могут давать один и тот же фенотип. Кроме того, ген определяли как единицу мутации. Эксперименты Бензера показали, что ген представляет собой линейную последовательность многих участков, в которых возможны разные мутации, и мы только что показали, как в комплементационных тестах можно выделять гены на основе происходящих в них мутаций. При этом ген понимается как последовательность, кодирующая синтез отдельной полипептидной цепи, и это представление основано на концепции Бидла и Тэй-тема «один ген — один фермент». Гены они определяли и как единицы рекомбинаций, хотя сейчас известно, что гены не представляют собой неделимые «бусины» на цепи, а рекомбинации происходят и внутри генов. Это и следовало ожидать, если предположить, что ген представляет собой всего лишь участок ДНК, любые нуклеотидные пары которой могут изменяться, в результате мутации и рекомбинаций.
В свете последних исследований, особенно секве-нирования (определения последовательности ДНК), приходится по-новому прдходить к вопросу о том что представляет собой ген. Так, оказалось, что в ДНК эукариот последовательности, кодирующие синтез белков, прерываются некодирующими последовательностями, называемыми интронами, которые удаляются непосредственно перед синтезом белка. Иногда на протяжении одного участка ДНК кодирующие последовательности, прерываемые интронами, сочетаются по-разному и кодируют разные белки. Если отождествлять отдельный ген с производством отдельного белка, то приходится признать, что одна и та же последовательность ДНК в таких случаях содержит несколько генов. Это только одна из трудностей. Другая состоит в том, что экспрессию, или «включенность», генов контролируют последовательности на участках ДНК, примыкающих к кодирующей последовательности, но не входящих в нее. Мутации в контролирующих участках могут привести к утрате геном функции, точно так же как и мутации внутри кодирующей последовательности. Поэтому, если выделять ген по критерию мутации, приходится признать, что контролирующие участки тоже относятся к гену. И наконец, подробный анализ ДНК-последовательностей целых геномов, включая и геном человека, предоставляют возможность опознать гены (по крайней мере, нечто вроде генов) на основании последовательности, а не мутаций. Белки со схожими функциями даже в очень отличающихся друг от друга организмах имеют много общего в строении. В настоящее время собраны обширные базы данных о ДНК-последовательностях, кодирующих белки; компьютерные программы могут просматривать все вновь определяемые последовательности и устанавливать возможные гены, предположительно кодирующие белки с теми или иными функциями. Даже если новая последовательность оказывается совсем не похожей на те, что уже имеются в базе, ученые все равно могут сделать вывод, что это ген, на основании хорошо известных признаков, общих для всех генов. Исходя из самого поверхностного анализа человеческого генома возможно предположить, что он содержит 30 000—50 000 генов, но если одна последовательность может включать более одного гена, то количество генов будет гораздо больше.
Генетические эксперименты Бензера и других ученых помогли составить представление о строении гена. Однако для любой науки характерно, что очередное открытие в отдельной области или технологии способно изменить основные ее положения. Для того чтобы функция генов стала более понятной, прочтите гл. 9, в которой более подробно объясняется, каким образом код ДНК преобразуется в структуру белка. Но прежде мы перенесемся через несколько лет и расскажем о другой процедуре составления карт, основанной на современном биохимическом анализе ДНК.








