 2015-06-28
2015-06-28 474
474ния ■ А между разными точками молекулы)
Рис. 18. Схема отрезка двухцепочной молекулы ДНК (по С. М. Гершензону)
положение пуриновых и пиримидиновых оснований нуклеотидов вдоль цепи ДНК очень изменчиво и характерно для каждого данного типа молекул ДНК. Значит, наследственная информация зашифрована различной последовательностью оснований.
Нуклеотидный состав ДНК значительно варьирует в зависимости от принадлежности организма к той или иной систематической группе (табл. 7). Специфичность ДНК выражается соотношением А + Т/Г + Ц, получившим название коэффициента видовой специфичности.
| 7. Нуклеотидный состав ДНК из различных (по А. Ленинджеру, 1976) | ИСТОЧНИКОВ | ||||||
| Нуклеотидный состав, | мол. % | Коэффициент | |||||
| Объект | А | Г | Ц | т | специфичности А + Т/Г + Ц | ||
30,9 19,9 19,8 29,4 1,52
Человек Животные:
| 28,3 1,36 29,2 1,38 27.1 1,19 32,9 1,79 24,9 1,00 23,6 0,93 29.2 1,50 36.2 2,70 21,1 0,72 12.2 0,35 |
| 29,3 | 21,4 | 21,0 |
| 28,8 | 20,5 | 21,5 |
| 27,3 | 22,7 | 22,8 |
| 31,3 | 18,7 | 17,1 |
| 25,0 | 25,1 | 25,0 |
| 24,7 | 26,0 | 25,7 |
| 30,8 | 21,0 | 19,0 |
| 36,9 | 14,0 | 12,8 |
| 21,0 | 29,0 | 28,9 |
| 13,4 | 37,1 | 37,1 |
овиа
курица
Растения, грибы, зерна пшеницы Дрожжи
Aspergillus niger Бактерии:
Е. coli
Staphylococcus ayreus
Clostridium perfinngens
Bnicella abortus
Sarcina lutea
| топоизомераза |
 В ДНК животных наблюдается избыток А + Т по отношению к Г + Ц. У грибов и бактерий встречаются формы как богатые А + Т, так и с преобладанием Г + Ц, в то же время есть близкие по коэффициенту специфичности к животным. Это говорит о том, что изменчивость в расположении оснований уже достаточна для того, чтобы обеспечить различия между генами этих организмов.
В ДНК животных наблюдается избыток А + Т по отношению к Г + Ц. У грибов и бактерий встречаются формы как богатые А + Т, так и с преобладанием Г + Ц, в то же время есть близкие по коэффициенту специфичности к животным. Это говорит о том, что изменчивость в расположении оснований уже достаточна для того, чтобы обеспечить различия между генами этих организмов.
Молекулы ДНК состоят примерно из 2-Ю3— МО8 и более нуклеотидов и имеют большую относительную молекулярную массу.
Репликация (удвоение) ДНК. ДНК находится в хромосомах, и репликация ее происходит перед каждым удвоением хромосом и делением клетки. Дж. Уотсон и Ф. Крик предложили схему удвоения ДНК, согласно которой спиралевидная двухцепочная ДНК сначала раскручивается (расплетается) вдоль оси. При этом водородные связи между азотистыми основаниями рвутся и цепи расходятся. Одновременно к нуклеотидам каждой цепи пристраиваются комплементарные азотистые основания нуклеотидов второй цепи, где против аденина встает тимин, против тимина — аденин, против гуанина — цитозин и т. д., которые с помощью ферментов ДНК-полимераз связываются в новые полинуклеотидные цепи. В результате из одной образуются две новые дочерние молекулы ДНК. Каждая дочерняя молекула, наследуя структуру одной цепи материнской молекулы, строго сохраняет специфичность заключенной в ней информации. Поскольку матрицей для репликации служит одна из двух цепей молекулы, такой тип синтеза ДНК носит название полуконсервативной ауторепродукции.
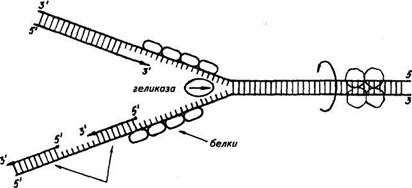
Дальнейшие исследования показали, что репликация бактериальных и других молекул ДНК начинается в определенной точке старта. В хромосомах эукариот обнаружено по нескольку таких начальных точек. Цепи ДНК в точке инициации репликации разъединяются под влиянием особого белка геликазы (рис. 19). Возникают одноцепочные участки ДНК, которые становятся матрицами для репликации-притяжения комплементарных нуклеотидов. Эти одноцепочные участки связываются с особыми белками, которые их стабилизируют (препятствуют их комплементарному взаимодействию). Особый фермент топоизомераза (у прокариот назьгаается ДНК-гиразой) способствует расщеплению спирали ДНК в области репликационной вилки.
Репликация на материнской цепи, идущей от точки старта в направлении 5'-*3', идет в виде сплошной линии. Эта цепь получила название лидирующей. Синтез на второй цепи 3'->5' идет отдельными фрагментами в противоположном направлении (тоже 5'-»3')- Эта цепь получила название запаздывающей. Фрагментами являются небольшие участки ДНК (у кишечной палочки около 2000 нуклеотидов, у эукариот около 200). Они называются по








