2015-06-26
2015-06-26 1607
1607При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития они проходят стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. X. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. В 1970 г. X. Темин и С. Мизутани, а также независимо от них Д. Балтимор открыли такой фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название ревертаза (обратная транскриптаза).
Наиболее детально изучена ревертаза ретровирусов птиц. Каждый вирион содержит около 50 молекул этого фермента. Обратная транскриптаза состоит из двух субъединиц - ά (65 кДа) и β (95 кДа), присутствующих в эквимолярном количестве. ά -Субъединица представляет собой N-концевую часть (две трети) β-субъединицы.
Обратная транскриптаза обладает по крайней мере тремя ферментативными активностями:
· ДНК-полимеразной, использующей в качестве матрицы как РНК, так и ДНК;
· активностью РНКазы Н, гидролизующей РНК в составе гибрида РНК-ДНК, но не одно- или двухцепочечную РНК;
· ДНК-эндонуклеазной.
Первые две активности необходимы для синтеза вирусной ДНК, а эндонуклеаза, по-видимому, важна для интеграции вирусной ДНК в геном клетки-хозяина. β-Субъединица ревертазы обладает всеми тремя активностями, в то время как ά-субъединица - только полимеразной и РНКазы Н.
Очищенная обратная транскриптаза синтезирует ДНК как на РНК-, так и на ДНК-матрицах. Чтобы начать синтез, ревертазе, как и другим полимеразам, необходим короткий двухцепочечный участок — праймер. Праймером может служить одноцепочечный сегмент как РНК, так и ДНК, которые в процессе реакции оказываются ковалентно связанными с новосинтезированной цепью ДНК.
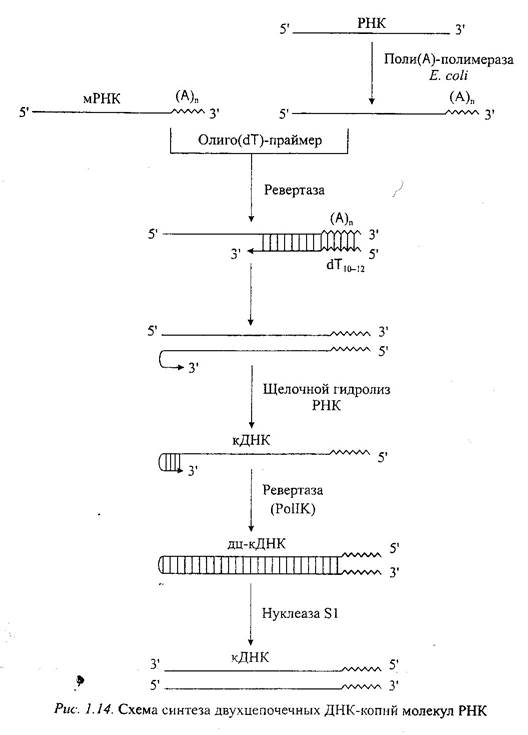
Обратную транскриптазу преимущественно используют для транскрипции матричной РНК в комплементарную ДНК (кДНК). Реакцию обратной транскрипции проводят в присутствии сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А)-содержащих мРНК используют олигo(dT) (рис.), а для молекул РНК, не имеющих 3'-поли (А)-концов, - химически синтезированные олигонуклеотнды, комплементарные 3'-концу изучаемой РНК. Кроме того, последний тип молекул РНК можно перевести в поли(А)-содержащие с помощью поли(А)-полимеразы Е. coli.
После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК.
По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. coli. Полученную двухцепочечную кДНК можно затем встраивать в клонирующие векторы, размножать в составе гибридных молекул ДНК и использовать для дальнейших исследований.