 2015-07-14
2015-07-14 904
904Весьма существенную роль в регуляции кислотно-щелочного баланса играют обменные процессы, происходящие в тканях, особенно в печени, почках, мышцах. Органические кислоты, могут подвергаться окислению с образованием либо летучих, легко выделяющихся из организма кислот (главным образом углекислоты), либо превращаясь в некислые вещества. Они могут соединяться с продуктами белкового обмена, полностью или частично утрачивая свои кислые свойства (например, соединение бензойной кислоты с глицином); молочная кислота, в больших количествах образующаяся при усиленной мышечной работе,
Ресинтезируется в гликоген, кетоновые тела - в высшие жирные кислоты и затем в жиры и т. д. Неорганические кислоты могут быть нейтрализованы солями калия, натрия, освобождающимися при дезаминировании аминокислот аммиаком с образованием аммонийных солей и т. д. Щелочи нейтрализуются главным образом молочной кислотой, которая при сдвиге активной реакции тканей в щелочную сторону усиленно образуется из гликогена. Кислотно-щелочной гомеостаз поддерживает и ряд физико-химических процессов: растворение сильных кислот и щелочей в средах с низкой диэлектрической постоянной (например, в липидах), связывание кислот и щелочей различными органическими веществами в недиссоциированные и нерастворимые соли, обмен ионов между клетками различных тканей и кровью и др.
Отмечая важность рассмотренных выше механизмов поддержания кислотно-щелочного гомеостаза, следует признать, что в конечном итоге узловым звеном в рассматриваемой гомеостатической системе является клеточный обмен, так как передвижение анионов и катионов между вне- и внутриклеточными секторами н их распределение в этих секторах являются прежде всего результатом деятельности клеток и подчинены потребностям этой деятельности.
Механизмы, обеспечивающие этот обмен, весьма разнообразны. Передвижение ионов зависит от градиента осмотического давления, проницаемости мембран, определяется динамическим электрическим потенциалом мембран и т. п.
Физиологические гомеостатические механизмы
Второй эшелон поддержания кислотно-щелочного гомеостаза представлен физиологическими регуляторными механизмами, среди которых главная роль принадлежит легким и почкам.
Благодаря буферам крови органические кислоты, образующиеся в процессе обмена, или кислоты, введенные в организм извне, не изменяют реакции крови, а лишь вытесняют углекислоту из ее соединения с основаниями; избыток же углекислого газа выводится легкими. Высокая диффузионная способность углекислого газа обеспечивает быстрое прохождение газа через мембраны и выведение его из организма. Скорость диффузии любого газа обратно пропорциональна квадратному корню из его молекулярной массы, а количество диффундирующего газа пропорционально его растворимости в жидкости.
Объединение этих двух законов диффузии позволяет сделать вывод, что углекислый газ диффундирует примерно в 20 раз интенсивнее кислорода:
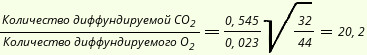
где 0,545 и 0,023 - коэффициенты растворимости соответственно СО2 и О2 в воде при t=38°C. Переход углекислого газа из крови в альвеолярный воздух объясняется имеющимся здесь градиентом Рсо2. Облегчается этот процесс двумя механизмами: переходом Нb в НbО2, вытесняющий как более сильная кислота углекислый газ из крови, и действием угольной ангидразы, которой принадлежит большая роль в освобождении свободной углекислоты в легких. Количество углекислого газа, выводимого из легких, зависит прежде всего от амплитуды и частоты дыхательных движений. Параметры дыхания регулируются в зависимости от содержания углекислоты в организме. В целом же отношение между Рсо2 в крови и легочной вентиляцией выражается следующим образом (Рут Г., 1978):
| Pco2 | = | P ·22,26· | CO2-продукция |
| Альвеолярная вентиляция |
где Рсо2 и Р (барометрическое давление) выражаются в миллиметрах ртутного столба, продукция СО2 - в молях, а альвеолярная вентиляция - в литрах.
Роль почек в поддержании кислотно-щелочного гомеостаза определяется главным образом их кислото -выделительной функцией. В физиологических условиях почки выделяют кислую мочу, pH которой колеблется от 5,0 до 7,0. Величина pH мочи может достигать 4,5, и, следовательно, концентрация свободных ионов Н+может в 800 раз превышать содержание их в плазме крови. Подкисление мочи в проксимальных и дистальных канальцах является результатом секреции ионов Н+, в образовании и секреции которых (ацидогенезе) важная роль принадлежит ферменту карбоангидразе (КА), содержащемуся в клетках канальцев. Фермент ускоряет достижение равновесия между медленной реакцией гидратации и дегидратации угольной кислоты (Н2СО3):
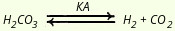
Скорость этой некатализируемой реакции возрастает при снижении pH. Ацидогенез обеспечивает выведение кислых компонентов фосфатного буфера (в процессе образования кислой мочи происходит превращение: НРО2-4 + H+ ---> H2PO4), а также слабых органических кислот: молочной, лимонной, β-оксимасляной и др. Процесс выделения эпителием почечных канальцев Н+ происходит против электрохимического градиента с затратой большого количества энергии и требует одновременной реабсорбции эквивалентного количества ионов Na+. Уменьшение реабсорбции натрия, как правило, сопровождается снижением ацидогенеза. Реабсорбированный в результате ацидогенеза Na+ образует в крови вместе с НСО-3 выделяемым из эпителия почечных канальцев, бикарбонат натрия. Ионы Н+, секретируемые клетками почечных канальцев, вступают во взаимодействие с анионами буферных соединений. Ацидогенез обеспечивает выделение преимущественно анионов карбонатного и фосфатного буферов, а также анионов слабых органических кислот.
При фильтрации соединений, содержащих анионы сильных органических и неорганических кислот (Cl-, SO2-4), в почках включается другой механизм - аммониогенез, обеспечивающий выведение кислот и предохраняющий от снижения pH мочи ниже критического уровня (рис. 39). Аммониогенез происходит на уровне дистальных канальцев и собирательных трубок. Образующийся в эпителии почечных канальцев NH3 поступает в просвет канальцев, где взаимодействует с Н+, происходящими от ацидогенеза. Таким образом, NH3 обеспечивает, с одной стороны, связывание Н+, а с другой - выведение анионов сильных кислот в виде аммонийных солей, в составе которых ионы Н+ не оказывают повреждающего воздействия на эпителий канальцев. Источником аммония является в основном глутамин крови. Около 60% NH3 образуется из глутамина путем его дезаминирования иод воздействием фермента глутаминазы I. Остальные 40% аммиака, образуются из других аминокислот.
Так как аммониогенез тесно связан с ацидогенезом, очевидно, что концентрация аммония в моче находится в прямой зависимости от концентрации в ней Н+. Закисление крови, приводящее к снижению pH канальцевой жидкости, способствует диффузии аммиака из клеток. Интенсивность экскреции аммония определяется также скоростью его продукции и скоростью потока мочи, от которой зависит время контакта между канальцевой жидкостью и эпителием почечного канальца, и, следовательно, своевременное удаление образующегося иона из клетки.
В регуляции экскреции почками кислот важную роль выполняют хлориды. В частности, увеличение реабсорбции НСО-3, как правило, сопровождается увеличением реабсорбции хлоридов. Ион С1- в общем пассивно следует за катионом Na+. Повышение концентрации гидрокарбонатов НСО-3 в моче обычно сопровождается уменьшением содержания в них хлоридов таким образом, что сумма этих анионов оказывается эквивалентной количеству Na+. Изменение транспорта хлоридов является следствием первичного изменения секреции ионов Н+ и реабсорбции бикарбоната и обусловлено необходимостью поддержания электронейтральности канальцевой мочи. Согласно другой точке зрения, первично изменяется транспорт хлоридов.
Кроме механизмов ацидо- и аммониогенеза, в сохранении иона Na+при закислении крови существенная роль принадлежит секреции ионов К+. Калий, освобождающийся из клеток при снижении pH крови, экскретируется почечными канальцами в повышенном количестве; одновременно при этом происходит увеличенная реабсорбция Na+. Этот обмен регулируется минералокортикоидами (альдостерон, дезоксикортикостерон). В нормальных условиях почки выделяют преимущественно кислые продукты обмена. При увеличении поступления в организм оснований реакция мочи становится более щелочной вследствие усиленного выделения бикарбоната и основного фосфата.
Определенное место в выделительной регуляции кислотно-щелочного гомеостаза занимает желудочно-кишечный тракт. Клетки слизистой оболочки желудка секретируют НСl, образующую из ионов Сl-, поступающих из крови, и ионов Н+, происходящих из желудочного эпителия. В обмен на хлориды в кровь в процессе желудочной секреции поступает бикарбонат. Защелачивания крови, однако, при этом не происходит, так как ионы Сl- желудочного сока вновь всасываются в кровь в кишечнике. Эпителий слизистой оболочки кишечника секретирует щелочной сок, богатый бикарбонатами. При этом ионы Н+ переходят в кровь в виде НС1. Кратковременный сдвиг реакции сразу же уравновешивается обратным всасыванием бикарбоната в кишечнике. В то время как почки концентрируют и выделяют из организма главным образом Н+ и одновалентные катионы, кишечный тракт концентрирует и выделяет двухвалентные щелочные ионы. При кислой диете увеличивается выделение главным образом двухвалентных Са2+ и Mg2+, при щелочной - выделение всех катионов.








