 2015-08-21
2015-08-21 1018
1018Р. Кох в 1876 г. доказал, что вегетативные клетки сибиреязвенного микроба обладают способностью формировать споры, в 1888 г. Серафини обнаружил капсулу микроба.
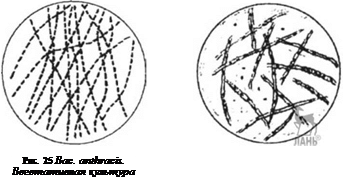
Морфология. Бациллы антракса довольно крупные (1-1,3 х 3,0-10,0 мкм) палочки, неподвижные, грамположительные; образуют капсулу и споры (рис. 25-28). Микроб встречается в двух формах: вегетативной, в виде палочки различной величины клеток (капсульных и бескапсульных), и споровой. Споры могут быть заключены в хорошо выраженный экзоспориум и находиться внутри и вне палочек в виде изолированных телец.
| Рис. 26 Споры |
 |
В окрашенных препаратах из крови и тканей больных или погибших от сибирской язвы животных бациллы располагаются одиночно, попарно и в виде коротких цепочек (3-4 клетки, окруженные капсулой). Концы палочек в цепочках прямые, с резко обрубленными концами, а свободные — слегка
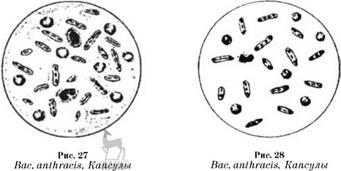
загругленные. Иногда цепочки имеют форму бамбуковой трости. В мазках из культур, на плотных и в жидких питательных средах палочки располагаются длинными цепочками.
В организме или при культивировании на искусственных питательных средах с большим содержанием нативного белка сибиреязвенная бацилла образует капсулу.
Сибиреязвенная бацилла во внешней среде при неблагоприятных условиях существования формирует споры. В каждой вегетативной клетке образуется только одна эндоспора, чаще располагающаяся центрально, реже — субтерминально. Споры овальные, иногда округлые. Размеры зрелых спор колеблются в пределах 1,2-1,5 мкм в длину и 0,8-1,0 мкм в поперечнике.
Культивирование. Сибиреязвенный микроб по способу дыхания относят к факультативным анаэробам: он хорошо размножается в обычных атмосферных условиях и в условиях пониженного содержания кислорода.
Бацилла антракса нетребовательна к условиям питания и хорошо растет на универсальных средах (МПБ, МПА, МПЖ, картофеле, молоке). Кроме того, может расти на различных растительных субстратах: настоях соломы, сена, экстрактах гороха, сои, вики, ломтиках вареного картофеля, свеклы, моркови и др.
Оптимальная температура роста культуры 35-37°С; при температуре ниже 12 и выше 45°С она не растет; оптимум рН среды 7,2-7,6.
На поверхности МПА в аэробных условиях при 37°С первые признаки роста появляются уже через 6-8 ч после посева,
17-24-часовые культуры имеют вид серовато-беловатых колоний с неровными краями и шероховатой поверхностью диаметром 3-5 мм. От их краев отходят завитки. Под лупой или малым увеличением микроскопа колонии имеют локонообразную структуру, что характерно для типичных вирулентных штаммов, образующих R-форму.
В МПБ и других жидких средах сибиреязвенная бацилла ^К-форма) через 16-24 ч образует на дне пробирки рыхлый белый осадок, сам бульон остается прозрачным, при встряхивании осадок разбивается на мелкие хлопья. Некоторые штаммы растут в виде нежных мелких хлопьев, взвешенных в столбике бульона, которые через 48 ч оседают на дно. Отдельные штаммы на 3-4-е сутки дают рыхлое пристеночное кольцо, пленка на поверхности среды не образуется.
Весьма характерный рост отмечают в столбике желатина при посеве уколом. По ходу укола на 2-5-е сутки появляется серовато-белый стержень, от которого под прямым углом ради-ально отходят нежные боковые отростки — более длинные по мере приближения к поверхности среды и постепенно укорачивающиеся по направлению вниз. Такая культура напоминает елочку, перевернутую верхушкой вниз. Постепенно верхний слой желатина начинает разжижаться, принимая сначала форму воронки, затем мешочка.
В молоке В. anthracis размножается быстро, вырабатывает кислоту и через 2-4 сут оно свертывается с последующей пептонизацией сгустка. На картофеле образует обильный, сухой, серо-белый налет, иногда с кремовым оттенком.
Агаровые и бульонные культуры некоторых штаммов интенсивно окрашиваются в светло-коричневый цвет вследствие окисления тирозина.
Биохимические свойства. В. anthracis вырабатывает следующие ферменты: липазу, диастазу, протеазу, желатиназу, де-гидразу, цитохромоксидазу, пероксидазу, каталазу и др. Некоторые штаммы образуют сероводород, особенно это свойство проявляется в средах, богатых пептонами; выделяют аммиак. Ферментирует с образованием кислоты без газа глюкозу, мальтозу, медленно сахарозу, трегалозу, фруктозу и декстрин. На цредах с глицерином и салицином возможно слабое кислотооб-разование. Арабинозу, рамнозу, галактозу, маннозу, рафинозу, инулин, маннит, дульцит, сорбит, инозит не сбраживает. Утилизирует цитраты, образует ацетал-метилкарбинол и вследствие этого дает положительную реакцию Фогеса — Проскауэ-ра. Синтезирует лецитиназу и медленно коагулирует растворы желтка куриного яйца. Редуцирует метиленовый синий и восстанавливает нитраты в нитриты. Вырабатывает желатиназу, а также протеазу и достаточно быстро гидролизует желатин и свернутую сыворотку.
Токсинообразование. Бацилла антракса образует сложный экзотоксин, включающий три компонента (фактора): эдемато-генный фактор (EF), протективный антиген (РА) и летальный фактор (LF), или соответственно факторы I, II, III. Их синтезируют капсульные и бескапсульные варианты микроба. Эдемато-генный фактор представляет собой липопротеин, вызывает местную воспалительную реакцию — отек и разрушение тканей.
Протективный антиген — носитель защитных свойств, обладает выраженным иммуногенным действием. В чистом виде нетоксичен. Летальный фактор сам по себе нетоксичен, но в смеси со II фактором (РА) вызывает гибель белых крыс, мышей и морских свинок.
Протективный антиген и летальный фактор — гетерогенные в молекулярном отношении белки.
Все три компонента токсина составляют синергическую смесь, оказывающую одновременно эдематогенное и летальное действия, каждый из них обладает выраженной антигенной активностью и серологически активен.
Инвазивные свойства микроба обусловлены капсульным полипептидом d-глутаминовой кислоты и экзоферментами.
Антигенная структура. В состав антигенов бациллы антракса входят неиммуногенный соматический полисахаридный комплекс и капсульный глутаминполипептид. Полисахаридный антиген не создает иммунитета у животных и не определяет агрессивных функций бациллы: всегда присутствует как у вирулентных, так и у авирулентных штаммов.
В связи с тем что полисахарид тесно связан с телом бактериальной клетки, он получил название соматического антигена.
Сибиреязвенный соматический антиген очень часто обозначают буквой С, капсульный полипептид — буквой Р.
Капсульный антиген бациллы антракса представлен сложным полипептидом d-глутаминовой кислоты; его принимают за группо-специфическое вещество, так как он дает перекрестные серологические реакции с полипептидом В. subtilis, В. cereus и В. megaterium. Активными антигенами также являются все три компонента сибиреязвенного экзотоксина.
Устойчивость. Устойчивость и длительность выживания у вегетативных клеток и спор возбудителя сибирской язвы различны. Вегетативные формы относительно лабильны, споры обладают высокой резистентностью.
В невскрытом трупе вегетативная форма микроба в результате воздействия протеолитических ферментов разрушается уже в течение 2-2 сут, в зарытых трупах сохраняется до 4 сут, через 7 сут завершается лизис бактерий даже в костном мозге.
В желудочном соке при 38°С гибнет через 30 мин, в замороженном мясе при -15°С жизнеспособна 15 сут, в засоленном мясе — до 1,5 мес. Навозная жижа, смешанная с сибиреязвенной кровью, губительно действует на вегетативные клетки уже через 2-3 ч, однако споры остаются в ней вирулентными в течение многих месяцев и лет.
Споры в запаянных ампулах сохраняют жизнеспособность и вирулентность до 63 лет, а в почве — более 60 лет.
К воздействию различных химических веществ вегетативные клетки малоустойчивы. Спирт, эфир, 2%-й формалин, 5%-й фенол, 5-10%-й хлорамин, свежий 5%-й раствор хлорной извести, пероксид водорода разрушают их в течение 5 мин.
Для уничтожения споровой формы возбудителя необходима более длительная экспозиция. Этиловый спирт в концентрациях от 25% до абсолютного разрушает споры в течение 50 сут и более, 5%-й фенол и 5-10%-й раствор хлорамина— через несколько часов и даже суток, 2%-й раствор формалина — через 10-15 мин, 1%-й раствор пероксида водорода — через 1 ч, 10% -й раствор гидроксида натрия — через 2 ч.
На споры высушивание не оказывает губительного действия: сухой жар при 120-140°С убивает споры только через 2-3 ч, при 150°С — через 1 ч.
Вегетативные клетки малоустойчивы к высоким температурам. При нагревании до 50-55°С гибнут в течение 1 н, при 60°С — через 15 мин, при 75°С — через 1 мин, при кипячении — мгновенно. Они чувствительны к высушиванию, однако при медленном высушивании происходит спорообразование, и микроб не гибнет.
К низким температурам бактерии малочувствительны: при -10°С сохраняются 24 сут, при-24°С до 12 сут. Воздействие прямого солнечного света обезвреживает бактерии через несколько часов.
Возбудитель сибирской язвы проявляет высокую чувствительность к пенициллину, хлортетрациклину и левомицетину, а также к литическому действию лизоцима.
Патогенность. К возбудителю сибирской язвы восприимчивы все виды млекопитающих. В естественных условиях чаще болеют овцы, крупный рогатый скот, свиньи, лошади, реже — ослы и мулы. Чрезвычайно восприимчивы козы, буйволы, верблюды и северные олени. Сибирская язва у свиней протекает, как правило, хронически, с длительным бациллоносительством. Среди диких животных восприимчивы все травоядные. Известны случаи заболевания собак, волков, лисиц, песцов, среди птиц — уток и страусов.
Патогенез. Бацилла антракса обладает выраженной инва-ивностью и легко проникает через царапины кожных покровов или слизистых оболочек. Заражение животных происходит преимущественно алиментарным путем. Через поврежденную слизистую оболочку пищеварительного тракта микроб проникает в лимфатическую систему, а затем — в кровь, где фагоцитируется и разносится по всему организму, фиксируясь в элементах лимфоидно-макрофагальной системы, после чего снова мигрирует в кровь, обусловливая септицемию.
Размножаясь в организме, бацилла антракса синтезирует капсульный полипептид и выделяет экзотоксин. Капсульное вещество ингибирует опсонизацию, в то время как экзотоксин разрушает фагоциты, поражает центральную нервную систему, вызывает отек, гипергликемию и повышение активности щелочной фосфатазы.
В терминальной фазе процесса в крови снижается содержание кислорода до уровня, несовместимого с жизнью. Резко нарушается метаболизм, развивается вторичный шок и наступает гибель животных.
Возбудитель сибирской язвы может выделяться из организма с бронхиальной слизью, слюной, молоком, мочой и испражнениями.
Лабораторная диагностика. При подозрении на сибирскую язву воспрещается вскрывать трупы павших животных. Для лабораторного исследования чаще всего направляют ухо павшего животного или толстые нефиксированные мазки крови из надреза сосуда на предметном стекле. При вынужденном убое или подозрении на сибирскую язву во время вскрытия осторожно отбирают кусочки селезенки, печени, измененные лимфоузлы, от трупов свиней — кусочки отечных тканей в области глотки и заглоточные лимфоузлы. Материал должен быть свежим: в разложившихся тканях бацилла антракса подвергается лизису. Направляют также пробы почвы, фуража, воды, шерсти и кожевенно-мехового сырья; объектами для серологического исследования в реакции преципитации служат пробы кожевенно-мехового сырья и разложившиеся ткани.
Исследование проводят по обычной схеме: бактериоскопия мазков, выделение и изучение свойств чистой культуры, биопроба на лабораторных животных, при необходимости, серологические исследования — реакция преципитации и иммуно-флюоресцентный анализ.
Бактериоскопия: из патологического материала готовят мазки, часть которых красят по Граму и обязательно на н аличие капсулы — по Михину, Ребигеру, Ольту и др.
Важным диагностическим признаком является обнаружение типичных по морфологии капсульных палочек.
Посев на питательные среды: исходный материал засевают в МПБ и на МПА (рН 7,2-7,6), посевы инкубируют при 37°С в течение 18-24 ч; при отсутствии роста выдерживают в термостате еще 2 сут.
Культуры просматривают, определяют их типичность, готовят препараты, микроскопируют. В мазках из культур обнаруживают бескапсульные палочки, расположенные длинными цепочками, и споры.
Биологическая проба: заражают белых мышей, морских свинок, кроликов одновременно с посевом материала на питательные среды.
Белым мышам вводят подкожно в заднюю часть спины (по 0,1-0,2 мл), морским свинкам и кроликам — под кожу в область живота (по 0,5-1,0 мл). Мыши погибают через 1-2 сут, морские свинки и кролики — через 2-4 сут. Павших животных вскрывают, делают мазки и посевы из крови, сердца, селезенки, печени и инфильтрата на месте инъекции исследуемого материала.
Идентификация бациллы антракса. В природе существует несколько видов аэробных споровых сибиреязвенноподобных сапрофитов: В. cereus, В. megaterium, В. mycoides и B.subtilis.
Так как они по морфологии и культуральным признакам во многом сходны с бациллой антракса, то часто при лабораторной диагностике возникает необходимость решить вопрос: выделена ли культура возбудителя сибирской язвы или подобного ему сапрофита?
Идентификацию и дифференциацию культуры проводят на основании главных и дополнительных признаков.
К главным относят патогенность, капсулообразование, определение подвижности, тест «жемчужного ожерелья», пробу со специфическим фагом, иммунофлюоресцентный тест,
к дополнительным — тесты на отсутствие гемолиза и лицетиназной активности, образование фосфатазы.
Бацилла антракса патогенна для лабораторных животных. Сибиреязвенноподобные сапрофиты не вызывают их гибель, за исключением В. cereus, которая убивает белых мышей при внут-рибрюшинном заражении. Массивную с четкими контурами капсулу в организме образует только возбудитель сибирской язвы.
Тест «жемчужногоожерелья», предложенный в 1953 г. Иенсеном и Клеемейером, основан на способности пенициллина угнетать синтез клеточной стенки бациллы антракса и образовывать сферопласты. Испытуемую трехчасовую бульонную культуру высевают на МПА в чашки Петри: в первой чашке содержится 0,5, во второй — 0,05 ЕД пенициллина в 1 мл среды, третья — контрольная. Посевы инкубируют 3 ч при 37°С.
На агаре с пенициллином бацилла антракса растет в виде цепочек, состоящих из шарообразных клеток, напоминающих ожерелье из жемчуга. Сибиреязвенноподобные сапрофиты на агаре с пенициллином этого феномена не образуют.
Проба с бактериофагом (лизабельность фагом): сибиреязвенный фаг, взаимодействуя с гомологичной культурой, вызывает ее лизис.
Эту высокоспецифичную реакцию применяют для идентификации бациллы антракса, а также дифференциации ее oт ложносибиреязвенных бацилл.
В качестве индикатора в России выпускают фаг ВНИИВВиМ, гамма-фаг МВА.
Иммунофлюоресцентный тест: идентификация возбудителя сибирской язвы при помощи флюоресцирующих антител — ориентировочный метод, который требует дополнительного изучения вирулентности, капсулообразования, фагочувствительности.
Подвижность устанавливается микроскопически или путем посева культуры уколом в столбик 0,3%-го МПА.
Неподвижные культуры растут только по ходу укола, подвижные — дают диффузный рост. Возбудитель сибирской язвы неподвижен, тогда как многие спорообразующие аэробные сапрофиты подвижны.
Гемолитическая активность не может быть надежным критерием дифференциации: как правило, бациллы антракса не гемолизируют эритроциты барана или же лизируют их очень медленно и незначительно, но этот признак у разных штаммов вариабелен.
Лецитиназная активность у возбудителя сибирской язвы низкая: медленно свертывает или вообще не свертывает желток куриного яйца. В. cereus интенсивно синтезирует лецити-назу и вызывает свертывание желтка через 6-10 ч. Не образует бацилла антракса и фосфатазу, в то время как сапрофитные споровые аэробы ее продуцируют.
Серологическое исследование: для обнаружения сибиреязвенных антигенов применяют реакцию преципитации по Асколи.
Эту реакцию используют для исследования на сибирскую язву кожевенного и мехового сырья, загнившего патологического материала, в котором происходит лизис бациллы антракса, а также для исследования свежего патологического материала и серологической идентификации выделенных культур.
РП по Асколи — достоверный и широко применяемый в практике тест серологической диагностики сибирской язвы. В качестве серологического теста, главным образом для изучения антигенного спектра бациллы антракса, применяют реакцию диффузионной преципитации (РДП).
Для идентификации и дифференциации возбудителя сибирской язвы от сходных микроорганизмов применяют селективную агаровую среду для выявления щелочной фосфатазы по изменению цвета колоний после воздействия паров аммиака. При этом сибиреязвенные колонии цвет не изменяют. С целью дифференциации от антракоидов рекомендована реакция диск преципитации, которая сочетает бактериологическую и серологическую диагностику.
Для выявления свежих случаев и ретроспективной диагностики сибирской язвы у животных предложен аллерген ВНИИВВиМ.
Иммунитет и средства специфической профилактики. Соматические полисахариды и капсульные полипептиды глу-таминовой кислоты бациллы антракса не способны обусловить синтез защитных антител. Эту функцию у бациллы антракса выполняет протективный антиген: будучи одним из факторов патогенности, он обусловливает формирование иммунитета к этой инфекции по типу антитоксического.
В настоящее время защитные антитела обнаруживают при помощи РСК, РДП и непрямого варианта метода флюоресцирующих антител в сыворотках животных, вакцинированных сибиреязвенным протективным антигеном или живыми споровыми вакцинами.
Переболевание животного сибирской язвой или же его вакцинация сопровождаются развитием гиперчувствительности замедленного типа. В результате естественного заражения и переболевания сибирской язвой у животных возникает длительный иммунитет.
Активная защита животных от сибирской язвы путем вакцинации — надежное средство профилактики данного заболевания. С этой целью применяют живые споровые сибиреязвенные вакцины.
Н. Н. Гинсбург в 1940 г. селекционировал из культуры вирулентного штамма вакцинный бескапсульный мутант СТИ-1. С 1942 г. вакцину СТИ, приготовленную из этого варианта, используют для профилактики сибирской язвы животных. Она зарекомендовала себя как высокоиммунный препарат. В настоящее время вакцину СТИ в споровой форме применяют для вакцинации животных против сибирской язвы. Иммунитет наступает через 10 сут и длится не менее 12 мес.
Новую вакцину против сибирской язвы из бескапсульного авирулентного штамма 55 выпускают в жидком и лиофилизированном виде. Вакцину вводят животным, начиная с 3-месячного возраста, однократно подкожно. Иммунитет наступает через 10 сут и сохраняется около 1 года.
В практике используют также живую вакцину ВГНКИ, ассоциированную живую вакцину против сибирской язвы и эмфизематозного карбункула крупного рогатого скота.
Для лечения и пассивной профилактики применяют антибиотики, противосибиреязвенную гипериммунную сыворотку и гамма-глобулин. Пассивный иммунитет наступает через несколько часов и сохраняется до 14 сут.
Использованная литература.

