2015-08-21
2015-08-21 1900
1900Чрезвычайно интересной особенностью пата следует считать их необыкновенную геологическ долговечность. Современный вид Triops саш formis был обнаружен в ископаемом состоянш триасовых отложениях на территории соврем ных ГДР и ФРГ. Детальный анализ хорошо сох] пившихся многочисленных отпечатков рачков их конечностей показал, что триасовый щип вполне сходен с современными. При этом он к в водоемах такого же типа, какие и сейчас насе; ны щитнями. Современные африканские т щитней также были найдены в ископаемом сося нии, но в юрских и в более молодых отложеню
Мы не знаем других подобных случаев суп ствования современных видов в столь отдалеш от нас геологические периоды. Щитней можно полным правом называть «живыми ископаемым
Как можно представить себе неизменность га на протяжении 200 млн. лет, отделяющих нас триасового периода? Английский ученый Лов х е р с т предполагает, что это объясняется гл ным образом постоянным разносом покоящш яиц щитней на огромные пространства, благод) чему не создается изоляции отдельных поселен рачков. Однако следует учесть и другие не ма важные биологические особенности щитней, и вящие их в особое положение в борьбе за сущи вование. В период их активной жизни в насел мых ими водоемах нет ни их врагов, ни их кош рентов. К тому же естественный отбор действ; на активных рачков всего в течение каких-i будь двух недель на протяжении целого го; Все остальное время рачки пребывают в ста! покоящихся яиц. Совершенно очевидно, что i
■ивование видов щитней зависит в первую оче- щ от стойкости таких яиц. Если допустить, что В яйца приобрели свои замечательные свойства ■ за несколько миллионов лет до наших дней, Качки в активном состоянии не подвергаются ес-■венному отбору, то' становится понятной по-■пельная консервативность видов этих уди-Вельных ракообразных.
КОНХОСТРАКИ (CONCHOSTRACA)
IB мелких, иногда пересыхающих водоемах
■ средней, так и северной части нашей страны,
■нередко и высоко в горах обитают рачки, менее
■метные и не достигающие столь крупных разме-
■, как щитни. Они обычно окрашены в бурова-
щ розоватые или зеленоватые тона. Все тело
■ покрыто двустворчатой хитиновой раковин-
1:1. которая у большинства видов имеет концен-
■иеские полосы прироста, несколько напоми-
Вющие кольца на раковинах двустворчатых мол-
Ikkob, например беззубки и перловицы. Эти по-
■и у рачков возникают после каждой линьки,
■кольку при линьке наружная часть старой ра-
■вины сохраняется, а затем раковина начинает
Ви по периферии. Число полос соответствует
■иу линек.
■ Обе створки скреплены одним поперечным силь-
■и мускулом-замыкателем. Рачки реагируют на
В\ость, сокращая этот мускул, причем створки
■овины плотно сжимаются и рачок, как мертвый,
иается неподвижно лежать на дне.
■Раскроем створки раковины, чтобы рассмотреть иоение тела рачка. Голова многих видов упло-■ена сверху вниз. Сидячие фасеточные глаза Вшжены и расположены на лобной ее части, а ■одного рода (Cyclestheria) даже соединяются ■кду собой. Перед ними находится простой нау-■шьный глазок. Передние антенны развиты ■о и не расчленены. Зато задние антенны длин-Re, двуветвистые, причем обе ветви снабжены ■гочисленными щетинками. Рачки плавают пре-Вщественно за счет взмахов задних антенн, дей-■ующих как весла. Лишь у очень обычного ро-Bynceus, способного широко раскрывать створ-Вшей шаровидной раковины, не только задние атенны, но и грудные ножки совершают весло-■разные движения.
■Число сегментов, составляющих грудной отдел, Иблется от 10 до 32, и соответственно этому ■меняется число пар грудных ножек. Они имеют ■иблизительно такое же строение, как у щитней,
■ вх размеры также постепенно уменьшаются по
■фавлению спереди назад. У самцов часть внут-
■енних лопастей первой, а иногда и второй пары
■дных ног преобразована в крючковидные при
|iinii, которыми самец удерживает самку при спа-
[ (рис. 193, Б). У самок наружные лопасти
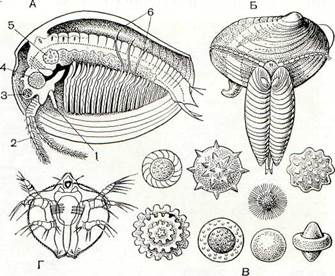
Рис. 193. Строение и биология Conchostraca:
А — строение самки Limnadia; Б — копуляция Cyzicus; В — яйца разных видов; Г — метанауплиус Lynceus; 1 — передняя антенна; 2 — задняя антенна; 3 — фасеточный глаз; 4 — мускул-замыкатель раковины; 5 — яичник; 6 — нитевидные отростки грудных ножек.
нескольких средних ног превращены в длинные, загнутые вверх нитевидные отростки, приспособленные для удерживания яиц на спинной стороне тела под раковиной. Однако основные функции грудных ног — это подача пищи вперед ко рту и дыхание. Жабрами служат ближайшие к основанию наружные лопасти всех грудных ног. Для обеспечения этих функций грудные ножки все время находятся в движении, благодаря чему вода между створками раковины беспрерывно обновляется. Тело заканчивается крупным твердым тельсоном, который либо сжат с боков и в таком случае снабжен парой крупных когтей, либо сплющен в спин-но-брюшном направлении и несет короткие коготки.
Большинство Conchostraca — донные животные. Наиболее крупные из их — Limnadia, достигающие длины 17 мм, лежат боком на дне. Cyzicus роется в грунте, взмучивая ил движением брюшка и отчасти ножек. Представители этих и других родов плавают лишь в период спаривания. Только сравнительно мелкий (4—5 мм) Lynceus brachy-urus, отличающийся шаровидной раковиной без полос прироста, обычно плавает спиной кверху, раскрыв створки раковины, и лишь изредка опускается на дно. Он питается главным образом планктонными организмами, в то время как Cyzicus ест грунт с находящимися в нем организмами, a Limnadia объедает микроскопические водоросли, находящиеся на дне и на поверхности водных растений. Крупные несъедобные частицы Limnadia склеивает специальным секретом, выделяемым же-
ПОДПИСИ К ЦВЕТНЫМ ТАБЛИЦАМ 49—56
| 51. Равноногие ракооб- Таблица54. Креветки: 1 — чилим широконосый травяной dalus latirostris); 2 — палемон изящный (Palaemon ele 3 — рак-щелкун альфеус Alpheus I ristata; 4 — шрнмс-медвежонок Sclerocranj salebrosa; 5 — леббеус Lebbeus sp.; С — леббеус гренландский (Lebh groenlandicus); 7 — пенеус японский (Penaeus ] cus); 8 — нотостомус Notostomus sp.; 9 — макробрахиум Macrobrachium 10, 11 — гипполите Hippolyte vari различно окрашенные; 12 — троглокарис Troglocaris anop mus; 13 — стенопус Stenopus hispidus, |
Таблица 49. Веслоногие ракообразные:
1 — сапфирина Sapphirina ovatolanceo-
lata;
2 — оитона Oithona plumifera;
3 — калянус Calanus finmarchicus;
4 — онцея Oncaea mediterranea;
5 — фенна Phoenna spinifera;
6 — калокалянус Calocalanus pavo;
7 — калокалянус оперенный
(С. plumulosus);
8 — корицеус Corycaeus obtusus;
9 — эухирелла Buchirella maxima;
10 — понтелла Pontella mediterranea;
11 — копилия Copilia vitrea;
12 — калянус Calanus plumchrus.
50. Усоногие ракооб-
Таблица
разные:
1 — морская уточка лепас Lepas anatife-
ra;
2 — морская уточка Dosima fascicularis;
3 — веррука Verruca stromia;
4 — Pollicipcs mitella;
5 — конходерма Conchoderma auritum
на коронуле Coronula diadema;
6 — Annandaleum japonicum;
7 — морской желудь балянус Balanus
improvisus;
8 — хирона Chirona evermanni;
9 — балянус Balanus balanus;
10 — хтамалюс Chtliamalus stellatus;
11 — ксенобалянус Xenobalanus globici-
pitis;
12 — криптолепас Cryptolepas rachianectis
Таблица
разные:
1,2 — гнатия зубчатая (Gnathia dentata), самец и самка;
3 — водяной ослик азелус (Asellus aquati-
cus);
4 — неофреатоикус несхожий (Neophrea-
toicus asslmilis);
5 — морской таракан мезидотея Mesidotea
entomon;
6 — ага Aega psora;
7 — циатура килевидная (Cyathura cari-
nata);
8 — гапломезус замечательный (Haplome-
sus insignis qrientalis);
9 — серолис Serolis sp.;
10 — батиномус гигантский (Bathynomus
giganteus);
11 — муннопсие типичный (Munnopsis ty-
pica);
12 — стортингура Storthyngura herculea;
13 — микрохарон Microcbaron sp.;
14 — порцеллио Porcellio scabpr;
15 — сферома зазубренная (Sphaeroma ser-
ratum);
16 — хемилепистус украшенный (Hemi-
lepistus cristatus);
17 — антарктурус ультраабиссальный
(Antarcturus ultraabyssalis).
Таблица 52. Разноногие ракообразные, или бокоплавы:
1 — брандтия Brandtia parasitica;
2 — нифаргоидес Niphargoides;
3 — аксельбекия Axelboeckia spinosa;
4 — эулимногаммарус пестрый (Eulim-
nogammarus virgatus);
5 — палласея двурогая (Pallasea bicor-
nis);
6 — паноплея двурогая (Panoploea bicor-
nis);
7 — акантогаммарус Годлиевского (Асап-
thogammarus godliewskii);
8 — леукотоэ Рихарда (Leucothoe ri-
cbiardii);
9 — грубия Grubia crassicornis;
10 — лизианасса Lysianassa punctata;
11 — бианколина Biancolina algicola;
12 — ампелиска Ampelisca diadema;
13 — цифокарис Рихарда (Cyphocaris
ricbardi);
14 — мегалуропус Megaluropus agilis;
15 — декзамине колючая (Dexamine spi-
nosa).
Таблица 55. Креветки:
1 — рак-щелкун альфеус Alpheus
vostokiensis;
2 — креветка-чистильщик на муре»
3 — рак-крот каллианасса (Gaily
sp.);
4 — ибакус ресничный (Ibacus oili
5 — глифокрангон Glyphocrangon
tacauda.
Таблица 56. Крабы:
1 — голубой краб-плавунец Callii
sapidus;
2 — пахиграпсус мраморный (Pachj
sus marmoratus);
3 — водорослевый краб пужетия Р
tia quadrilrons;
4 — травяной краб карцинус (Cat
maenas);
5 — хиас Hyas coarctatus;
6 — краб-стригун Chionoecetes opi!
7 — канцер Cancer pagurus;
8 — майя Maja squinado;
9 — калаппа Calappa sp.;
10 — рандаллия Randallia eburnea;
11 — лиреидус Lyreidus tridentatus;
12 — ламбрус Lambrus validus;
13 — дромия обыкновенная (Dromia
garis);
14 — краб-горошина пиннотерес Pini
res boninensis.
Таблица 53. Представители разных отрядов ракообразных:
1 — ветвистоусные рачки дафнии (Daph-
nia);
2 — усоногие раки морские желуди (Ba-
lanus);
3 — бокоплав морская козочка капрелла
(Caprella) с молодью.
 иезами ножек, и выорасывает через заднюю щель раковины.
иезами ножек, и выорасывает через заднюю щель раковины.
Размножение Conchostraca происходит по-разному. Все представители семейства Limnadiidae размножаются только партеногенетически, самцы среди них неизвести*ы. У других семейств самцы встречаются регулярно, причем, чем южнее оби-■ют рачки, тем больше относительная числен-■сть самцов. При спаривании самец удерживает ■мку крючковидными отростками задних антенн н просовывает задний конец тела в заднюю щель иежду створками раковины самки. Яйца отклады-ваются в пространство между спинной стороной на самки и ее раковиной. У Cyclestheria часть идних грудных сегментов расширена таким об-|зом, что перед ними образуется настоящая вы-■дковая сумка. Проносив некоторое время яйца на спине под раковиной, самка линяет, и яйца падают на дно водоема.
Эй яйца окружены плотной и прочной оболоч-*кой и способны переносить высыхание, замерзание и другие неблагоприятные условия. Нередко («ни имеют причудливую форму, у многих видов -они снабжены выростами, способствующими их переносу ветром и различными животными (рис. 193, В). Яйца некоторых видов сохраняют жизне-шособность в течение 15 лет.
После того как яйцо попадет в благоприятные ря его развития условия, из него выходит лишка, находящаяся на стадии науплиуса. Нау-шшус Conchostraca отличается очень маленькими передними антеннами. На следующей стадии — геанауплиуса — у личинки развивается зачаток. У большинства Conchostraca эта за-
чаточная раковина уже двустворчатая и очень напоминает раковину ветвистоусых ракообраз-шх(см. ниже). У Lynceus она имеет вид плоского щта,похожего на карапакс щитней (рис. 193, Г), только на более поздних стадиях развития она перегибается пополам и становится двустворчатой. Все развитие происходит очень быстро. Например, Cyzicus достигает половозрелости через 19 суток ше выхода личинки из яйца. Современные Conchostraca не имеют никакого практического значения. Однако вымершие виды № ракообразных привлекают к себе внимание геологов. В пресноводных отложениях, вообще пень бедных ископаемыми, хорошо сохраняющие-й раковинки Conchostraca встречаются довольно ито. По ним удается определять геологический ■раст соответствующих слоев. В предыдущие геологические периоды Conchost-иса были, по-видимому, более многочисленны и пре распространены, чем ныне. Вымерших видов ввестно примерно 300, а современных почти вдвое иьше. Наиболее древние Conchostraca найдены {девонских отложениях. Среди древних вымер-ю видов, особенно триасовых, многие жили в пре, где сейчас Conchostraca отсутствуют.
ПОДОТРЯД ВЕТВИСТОУСЫЕ РАКООБРАЗНЫЕ (CLADOCFRA)
Почти в каждом пресноводном водоеме, начиная с огромных глубоких озер и кончая очень мелкими водоемами, любой лов сачком или специальной сеткой из мельничного шелкового газа приносит десятки и сотни мелких рачков, часто называемых водяными блохами (табл. 48). Они оживленно снуют в толще воды, делая характерные взмахи задними антеннами, а некоторые из них передвигаются по дну или по подводным растениям. Их хорошо знают не только ученые-зоологи, но и все любители аквариумов, которые кормят ими, живыми или сушеными, своих рыбок. Ветвистоусых ракообразных знают и рыбоводы, занимающиеся их разведением для кормления молоди рыб, выращиваемой на рыбоводных заводах. Многие из этих рачков всю жизнь проводят в толще воды и составляют значительную часть пресноводного планктона. Иногда они развиваются в таком количестве, что вода приобретает буроватый оттенок. Наряду с планктонными есть и донные виды. Есть и такие, которые то прикрепляются к растениям или опускаются на дно, то всплывают в толщу воды. Подавляющее большинство видов ветвистоусых обитает в пресных водах, но есть и несколько морских видов. Размеры этих рачков незначительны, обычно меньше 1 мм, редко до 5 мм и в исключительных случаях до 18 мм. Поэтому для детального изучения их строения приходится пользоваться микроскопом.
Тело большинства ветвистоусых ракообразных, например дафнии (рис. 194), заключено в хитиновую полупрозрачную двустворчатую раковинку, а голова вытянута в направленный вниз клюв, или рострум. На лобной части головы расположен непарный фасеточный глаз, возникший в результате слияния двух боковых глаз, а перед ним — простой науплиальный глазок, отсутствующий у некоторых видов.
Число фасеток, образующих сложный глаз, различно у разных видов и зависит от способа их питания. В глазу отфильтровывающей мелкие частицы дафнии насчитывается всего 22 фасетки, а у хищных Evadne и Leptodora соответственно 80 и 300. При нападении на мелких животных глаза хищных ветвистоусых имеют важное значение. Под рострумом прикрепляются палочковидные, обыкновенно относительно очень маленькие передние антенны. Зато задние антенны непропорционально велики по сравнению с телом. Они двуветвисты, причем каждая ветвь снабжена длинными перистыми щетинками^ Задние антенны служат основным органом передвижения ветвистоусых. Одновременно взмахивая обеими задними антеннами, рачки отталкиваются ими от воды и таким образом плывут короткими скачками. На нижней поверхности головы находится
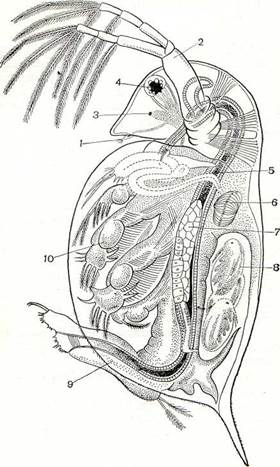
Рис. 194. Строение дафнии:
1 — передняя антенна; 2 — задняя антенна; 3 — науплиальный глазок; 4 — фасеточный глаз; 5 — кишечник; 6 — сердце; 7 — яичник; 8 — эмбрионы в выводковой сумке; 9 — брюшко; ю — грудная ношка.
рот, окруженный спереди верхней губой, с боков жвалами и сзади челюстями.
Грудной отдел ветвистоусых укорочен и, судя по числу пар ножек, состоит из 4—6 сегментов. Он, как уже указывалось, помещается внутри двустворчатой раковины, но у самок между спинной поверхностью тела и спинным краем раковины имеется обширная полость, выполняющая роль выводковой сумки. В эту сумку откладываются яйца, и при благоприятных условиях там же они развиваются. У хищных ветвистоусых раковина уменьшена в размерах, оставляет непокрытым почти все туловище и прикрепляется к его спинной стороне (табл. 48, 2, 5, б, 8, 14, 18); в таких случаях она служит только для вынашивания яиц.
Строение грудных ножек различно у разных видов и зависит от характера их питания. Эти ножки в общем напоминают листовидные ножки
жаброногов, щитней и Conchostraca. У болыш ства ветвистоусых ракообразных грудные но;я служат для отфильтровывания мелких взвеше ных в воде частиц, которыми рачки питании В соответствии с этим лопасти грудных нож снабжены многочисленными перистыми щетш ками, образующими вместе фильтрационный I парат. Ножки совершают до 300—500 взмая в 1 мин.
В простейшем случае, например у Sida (■ 195, 1), вода входит между створками в пере; ней части раковины. При одновременном дввд нии всех 6 пар ножек вперед между ними возш кает как бы ряд всасывательных камер, щ просачивается вода. При этом щетинки отфилы ровывают всю мелкую взвесь. Движение нош назад сжимает камеры и выгоняет воду чер заднюю часть раковины. Отфильтрованные 1 тицы направляются при помощи специальш щетинок в продольный желоб, расположенный! нижней стороне туловища между основания! грудных ножек. По этому желобу находящим на его стенках щетинками пищевой комок прота; кивается вперед, по направлению ко рту.
У Daphnia этот аппарат усложнен и усове) шенствован (рис. 195, 2). Из пяти пар ее грудин ножек фильтруют только третья и четвертая. Щ движении первой пары ножек вперед вода пост! пает в фильтрующую камеру. В это время трети и четвертая пары изгибаются вперед, обеспечив дальнейший ток воды. Отфильтрованные пищевн частицы поступают в брюшной желобок. Пят! пара ног ограничивает сзади фильтрующую кай ру. Делая взмах назад, она вместе с четвертой:! рой вызывает выводящий ток воды. Таким обр! зом, между грудными ножками дафнии функл разделены, что говорит об их большой специал зации.
Ближайшие к основанию наружные лопаа грудных ног фильтрующих ветвистоусых ракоо! разных превращены в жабры. По системе полости кровь поступает в жабры и обогащается в них к| лородом. В плазме крови ветвистоусых растворе гемоглобин, от которого в значительной стеши зависит окраска рачков. Его концентрация ш няется. Если содержание кислорода в воде ей кое, кровь приобретает темно-красный цвет, ее» кислорода в воде много, кровь окрашена значж тельно светлее. Для изменения окраски рачке требуется около 10 суток. Дафнии с темно-краш кровью в воде с низким содержанием кислород живут значительно дольше, чем дафнии, имеют» светлую, бедную гемоглобином кровь. Очевидна гемоглобин, как и у позвоночных животных, ев» зывает кислород.
Кровь приводится в движение мешковиден! сердцем, взбалтывающим ее в полости тела (pi 194, 6). Настоящих кровеносных сосудов ни, Сердце дафнии бьется очень часто. При темпер!
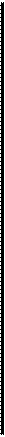 ре 10 °С оно делает 150, а при температуре 1°Сдаже 500 ударов в 1 мин. Вовсем иначе устроены и действуют грудные ига хищных ветвистоусых ракообразных юл. 48). Их боковые лопасти недоразвиты или вугствуют, жабр нет совсем, а хорошо разви-■е внутренние ветви снабжены сильными шипа-га и шиловидными щетинками. При помощи груд-1 ног рачки схватывают других мелких живот-1, служащих им пищей. Хищные ветвистоусые «принимают кислород скоплением железистых веток, находящихся на голове. [Задний брюшной отдел тела ветвистоусых уко-иен, часто изогнут под прямым углом и может ютаться между створками раковины. Он обыч-|заканчивается двумя когтями. Однако у мно- ш хищных видов задний отдел имеет форму вбки, сохраняет сегментацию и иногда продол-■ется в длинный шип, назначение которого ос-бется непонятным.
ре 10 °С оно делает 150, а при температуре 1°Сдаже 500 ударов в 1 мин. Вовсем иначе устроены и действуют грудные ига хищных ветвистоусых ракообразных юл. 48). Их боковые лопасти недоразвиты или вугствуют, жабр нет совсем, а хорошо разви-■е внутренние ветви снабжены сильными шипа-га и шиловидными щетинками. При помощи груд-1 ног рачки схватывают других мелких живот-1, служащих им пищей. Хищные ветвистоусые «принимают кислород скоплением железистых веток, находящихся на голове. [Задний брюшной отдел тела ветвистоусых уко-иен, часто изогнут под прямым углом и может ютаться между створками раковины. Он обыч-|заканчивается двумя когтями. Однако у мно- ш хищных видов задний отдел имеет форму вбки, сохраняет сегментацию и иногда продол-■ется в длинный шип, назначение которого ос-бется непонятным.
[Наблюдая за поведением ветвистоустых рачков [водоеме или в аквариуме, нетрудно убедиться;нш. насколько различны повадки разных ви- ш. Особенно многочисленны рачки, проводящие [в жизнь в толще воды. Планктонные дафнии ■некоторые другие ветвистоусые после взмаха ■ними антеннами и вызванного этим движением ■ика некоторое время неподвижно парят в воде, иагодаря тому что центр тяжести их тела нахо-вся ниже места прикрепления антенн, в момент 1ра антенн тело рачка наклоняется, а в период юя снова выпрямляется. Распростертые ан-препятствуют погружению рачка (рис.
Л
[ланктонные ветвистоусые из рода Bosmina Цот иначе. Они беспрерывно машут задними рами, причем передняя часть их тела при
«дом ударе несколько приподнимается. Одна-длинные хоботообразные передние антенны 1ат балансиром, возвращающим тело в гори-тальное положение (рис. 195, 4). Если босми-отрезать передние антенны, она начинает бес-(ощно кувыркаться и теряет способность к пла-
пю.
!реди зарослей подводных растений в прибреж-1части озер и прудов часто попадается круп-|(до 4 мм) ветвистоусый рачок Sida crystalline, ррый, как бы устав беспрерывно плавать, вре-р времени прикрепляется к растениям при по-| специальной присоски, находящейся на части его головы. Другой обитатель зарос-I- Simocephalus — цепляется за растения ще-1ами задних антенн. Маленький шарообраз- ixudopyc (Chydorus sphaericus), поплавав не-ie время, садится на водоросли и начинает [зать по ним, удерживаясь щетинками края 1вины и отталкиваясь второй парой грудных ■и брюшком. Наиболее своеобразен способ ювюкения Scapholeberis mucronata, который
благодаря присутствию несмачиваемых щетинок на нижнем крае раковины может прикрепляться снизу к поверхностной пленке натяжения воды и быстро скользить по ней спинной стороной вниз (рис. 196).
Наряду с такими постоянно или временно плавающими формами существует большое число видов, плавающих очень плохо и постоянно ползающих по дну. Некоторые из них способны даже зарываться в поверхностный слой ила.
С жизнью многих ветвистоусых ракообразных в толще воды связано одно любопытное явление,
Рис. 195. Движения ветвистоусых:
1,2 — токи воды (обозначены стрелками), вызываемые движениями конечностей Sida (1) и Daphnia (2); 3,4 — движения в воде Daphnia (5) и Bosmina {4).
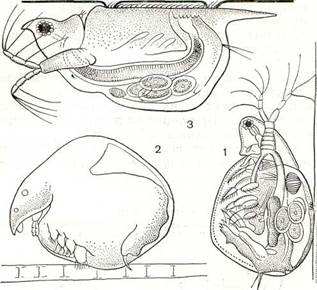
Рис. 196. Прикрепляющиеся ветвистоусые:
1 — Simocephalus vetulus на нити водоросли; 2 — Chydorus sphae-ricus на нити водоросли; 3 — Scapholeberis mucronata у поверхности воды.
получившее название цикломорфоза. Сущность его заключается в том, что форма и величина раковины определенных видов закономерно изменяются по сезонам года. В летнее время у некоторых видов дафнии на голове появляется так называемый «шлем», а у босмин на спинной стороне— «горб». Кроме того, у дафний удлиняется задний шип на раковине, а у босмин — передние антенны (рис. 197).
Эти изменения происходят параллельно изменению температуры воды. В данном случае важна, однако, не сама температура, а ее воздействие на плотность воды. Прогретая вода обладает меньшей плотностью, чем охлажденная. Для движения и парения в менее вязкой прогретой воде необходимы более длинные выросты, а иногда и иная форма тела. Поэтому летние формы дафний и босмин развивают специальные приспособления для передвижения и парения в таких условиях.
Большинство ветвистоусых ракообразных, как уже указывалось, питается, отфильтровывая мелкую, находящуюся в воде взвесь. Основной их пищей служат бактерии, одноклеточные водоросли и мелкие органические остатки ■— детрит. Наибольшее значение имеют бактерии. В опытах многих исследователей удавалось воспитывать ветвистоусых при кормлении их одними бактериями, а американский зоолог Банта получил в таких условиях около 1600 поколений дафний. В природных водоемах ветвистоусые прекрасно живут и размножаются при концентрации бактерий не менее 1 млн. в 1 см8 воды. Если численность
бактерий меньше 500 тыс. на 1 см3, наступлеи половозрелости дафний задерживается; если | численность бактерий падает ниже 200 тыс. I 1 см3, жизнь рачков становится невозможно; С другой стороны, избыточное количество баи рий (более 3—5 млн. на 1 см3) также вредно влия на рачков и вызывает прежде всего замедлен] или приостановку их размножения, а иногда да) их гибель. Это объясняется выделением бактерв ми продуктов обмена веществ, ядовитых для ве вистоусых.
От количества бактерий в водоеме зависит чи ленность ветвистоусых. Неоднократно наблкд лось уменьшение числа ветвистоусых в пери] массового развития сине-зеленых водорослей. В которые исследователи предполагали вредное т действие сине-зеленых на рачков. Однако, по м ным Е. Ф. Мануйловой, бурное.разил сине-зеленых само по себе никакого влияния рачков не оказывает. Оно подавляет развш бактерий и именно таким образом косвенно щ действует на ветвистоусых. Часто наблюдающий периоды резкого увеличения численности вети стоусых обычно следуют за периодами размно» ния бактерий.
Помимо бактерий, фильтрующие ветвистоуа используют в пищу одноклеточные водоросл Они могут заглатывать только самые мелкие вщ протококковых и зеленых водорослей, котор! обычно развиваются в большом количестве в пр; дах, но не в крупных озерах. Обычные, особен! колониальные, планктонные водоросли слипш велики для того, чтобы быть проглоченными ра1 ками. Во многих случаях водоросли приобретя! большое значение для ветвистоусых не потому, 4 поедаются ими, а потому, что после отмирал вызывают массовое развитие бактерий — осш ного корма рачков.
При фильтрации пищи рачки не могут отсорл ровывать съедобные частицы от несъедобна! Они способны только отбирать частицы по их щ меру, отбрасывая слишком крупные. С этим «заны, по наблюдениям В. М. Р ы л о в а, те и менения в составе планктона, которые наблга ются при проникновении планктона стоячих i доемов в текучие. Эти изменения сводятся глл ным образом к резкому уменьшению численном ветвистоусых ракообразных, в то время как ш ленность других планктонных животных, напр! мер коловраток, существенно не меняется. Ом зывается, что речной поток несет мелкие мин ральные частицы и попавшие в него планктошш ветвистоусые отфильтровывают их и заполню ими свой кишечник. Рачки идут ко дну и пощ бают, а коловратки, захватывающие пищу инш способом, и притом еще более мелкую, остаюи невредимыми.
Фильтрация пищи рачками происходит беспре рывно. Время, нужное для заполнения кшпечи
на, колеблется от 10 до 240 мин и зависит от ве-ичины рачков, размеров пищевых частиц, их концентрации и температуры. Мелкая босмина в культуре водоросли Ankistodesmus наполняет кишечник в течение 10 мин, а в культуре бактерии Bacillus coli тратит на это 2—2,5 ч. В течение су-юкодна дафния поглощает от 4,8 до 40,8 млн. бак-крий (в зависимости от их концентрации). Даже реле наполнения кишечника рачки продолжают фильтровать, и отфильтрованные частицы двигайся вперед по их брюшному желобку. Но в рот и не попадают: ротовые придатки и задний от-дел тела выталкивают их снова в воду. Рачки ыа-шают заглатывать пищу только после того, как ишечник хотя бы частично освободится от ее ос-татков.
IСовсем иначе добывают себе пищу хищные ви- ш, Самый крупный совершенно прозрачный вет-истоусый рачок Leptodora kindtii (табл. 48, 14) подвижно парит в воде и своим громадным гла-|зои высматривает добычу. Достаточно нескольких ильных взмахов его задних антенн для того, чтобы он настиг какого-нибудь более мелкого шантанного рачка или коловратку, схватил жертву далеко вынесенными вперед грудными [ногами и вонзил в нее острые зазубренные жвалы. Приблизительно так же охотится и другой крупный хищник — Bythotrephes (табл. 48, 2). Оба рачка сначала размельчают жертву жвалами, а ■тем всасывают жидкую пищу. Они нападают Ви на всех планктонных животных, включая ршенную молодь, но не трогают сидячих и даже ■временно прикрепляющихся рачков, например иду и симоцефалюса.
I Лептодора и битотрефес весьма прожорливы. Ври температуре 20—25 °С каждая лептодора ведает за сутки более 50, а битотрефес около IS рачков. Соответствующие расчеты показывают, но в некоторых водоемах, например в Рыбин-■еком водохранилище, хищные ветвистоусые вые- \щ заметную часть зоопланктона (более 40% вех планктонных животных). Наносимый ими |шш образом вред не так велик, поскольку они 1свою очередь интенсивно потребляются многими Вами.