 2015-08-21
2015-08-21 648
648Среди всех групп беспозвоночных животных тип членистоногих1 выделяется наибольшим разнообразием приспособлений к самым различным условиям существования, изумительным богатством форм и огромным числом видов. Число видов членистоногих приближается к 3 миллионам и намного превышает число видов всех остальных типов животных и растений, вместе взятых, включая микроорганизмы. Особенно обильны видами насекомые, на долю которых падает 90% известных видов членистоногих.
Именно тонкими приспособлениями к использованию всевозможных местообитаний, всяких источников пищи, часто недоступных для других животных, объясняется многообразней обилие видов членистоногих.
Трудно найти такие места, где не было бы членистоногих. Они живут во всех морях и океанах —• и в толще воды, и на дне, и в грунте на разных глубинах; они обитают и во всевозможных пресных водоемах — не только в реках и озерах, но и в мелких пересыхающих лужах, в подземных водах и пещерах, в скоплениях воды в дуплах деревьев. Некоторые виды приспособились к жизни в очень соленых водоемах, другие — в горячих источниках. Кажется, все разнообразие водной среды на поверхности нашей планеты освоено членистоногими.
Большая армия представителей этого типа приспособилась и к жизни вне воды — на суше, где членистоногие особенно многообразны. Весь слой почвы, пронизанный корнями растений, обильно заселен и членистоногими — во многих лесных и луговых почвах миллион особей клещей и ного-хвосток на 1 м2 не редкость. Огромное количество членистоногих обитает на поверхности различных частей растений и внутри растительных тканей.
Многие членистоногие — паразиты, постоя] или временные, наружные или внутренние, других видов членистоногих, так и животных носящихся к другим типам, причем, напри среди насекомых число видов паразитов не гим, вероятно, уступает числу остальных bi Иногда членистоногие столь изменяются под в нием паразитического образа жизни, что, перестают быть похожими на членистонога
Какими только способами, какими только ществами членистоногие не питаются. (фильтруют воду, потребляя мелкие планктс» организмы и мелкие взвешенные частицы ре гающихся остатков организмов; другие роют грунте или почве, пропуская через кише1 большое количество минеральных частиц, ч' усвоить рассеянные между ними органически! щества; есть членистоногие, которые питаюто кими трудноперевариваемыми веществами, рог и воск; есть существующие за счет клет ки. Некоторые способны усваивать с пома симбиотических микроорганизмов атмосфер азот. Огромное число представителей разных к сов членистоногих — хищники и паразиты; наземных членистоногих, особенно насеко.) один из основных источников пищи — ткани вых высших растений.
Во всех климатических зонах можно ветре1 членистоногих. Огромно их количество и вел многообразие в тропиках, но есть они и в вью' широтах Арктики и на побережье Антарктиды относительно недавно обнаружены новые i клещей и ногохвосток. Членистоногих мо встретить и в самых сухих пустынях. Обр1 можно сказать, что членистоногие — это тип£ вотных, который по полноте овладения по# ностыо земного шара не имеет равных.
По многим признакам строения членистой'1 близки к кольчатым червям, особенно к пол* там. От близких к ним форм когда-то давно,3 в те геологические эпохи, в отложениях которых* не сохранилось ископаемых остатков, отделов! предки современных членистоногих.
ноя нервная система у более примитив-аистоногих очень сходна с лестничной шстемой полихет.
ические признаки членистоногих свя-м, что у них тело покрыто очень плотной й кутикулой, которая нередко об-гень твердый панцирь. Кутикула у чле-[X выделяется наружным слоем клеток, темой гиподермой, имеет очень сложное и сложный химический состав. Раньше что у членистоногих кутикула состоит хитина. Теперь выяснилось, что, кроме состав ее входят многие другие соедине-говном задубленные белки, и что у боль-членистоногих на долю хитина падает з трети веществ, из которых состоит ку-Хитин — это сложный полисахарид, в шекулы которого входят атомы азота, астичен, прочен и устойчив к химиче-действиям. Нередко в кутикуле содер-ого карбоната кальция. гть и твердость кутикулы членистоно-лько велики, что в тех местах, где она большой толщины, покровы становятся 1. Подвижность сегментов тела друг ьно друга, обеспечивающая движение, тоногих достигается тем, что между участками (склеритами) сохраняются тые участки с очень тонкой кутикулой; эепонки на сгибах образуют складки, [ющиеся при растягивании тела. Далеко ■менты у членистоногих подвижны друг ьно друга, очень часто наблюдается лияние соседних сегментов, особенно х отделах тела.
членистоногих обычно можно различить — г о л о в у, на которой находятся органы чувств, служащие для ориенти-ространстве (глаза, усики и др.), и орга-ывания пищи. Сегменты головы у чле-х слитные. Далее следует грудь — тела, на котором располагаются основательные придатки — ноги, а у насе-крылья. Задний отдел тела называет-ш к о м. У некоторых членистоногих, у многоножек, а из ископаемых — у в) все туловищные сегменты более или ородны, и тогда в теле животного разли-ко 2 отдела — голову и туловище. У мно-[стоногих (у паукообразных, десятино-) головной и грудной отделы сливаются.)й вообще утрачиваются границы между тело слитное, (гаются членистоногие с помощью конеч-
>ку конечности у членистоногих, как и покрыты кутикулой, их движение возке только при разделении ног на чле-строению конечностей, па которые опи-
раются членистоногие при движении по твердой поверхности или с помощью которых они плавают, весь тип и получил свое название.
Некоторые конечности на голове членистоногих превращаются в ротовые органы — челюсти, использующиеся хотя бы частично для захватывания или размельчения пищи. Это тоже признак, характерный для типа членистоногих. Ротовые части бывают настолько видоизменены, что их трудно даже признать конечностями, например хоботок комара или клопа. Число пар ног, которые служат для передвижения, неодинаково у разных членистоногих. Для насекомых характерно развитие 3 пар двигательных конечностей, для паукообразных — 4 пар, а у некоторых многоножек число ног превышает сотню.
Нередко туловищные конечности служат не только как ноги, но и как жабры, присоски и другие органы.
У членистоногих двигательная мускулатура состоит из отдельных мышц, концы которых присоединяются к разным утолщенным участкам покровов, отделенным друг от друга мягкой перепонкой. Движение членистоногих обеспечивается не изгибами тела при сокращении всей мускулатуры туловища, как у червей, а сокращениями обычно немногих мышц, связанных с конечностями или другими двигательными придатками (крылья насекомых, вилочка ногохвосток и др.). Мышцы членистоногих обладают исключительно высокой способностью к сокращению. Все они поперечнополосатые, причем обнаруживаемая под микроскопом поперечная исчерченность мышечных волоконец у членистоногих выражена больше, чем у позвоночных,— мышцы членистоногих сильнее наших, если, конечно, учитывать размеры. Так как панцирь членистоногих служит и местом прикрепления двигательных мышц, он является и скелетом, называемым «наружным» в отличие от привычного нам внутреннего скелета позвоночных.
Панцирь членистоногих лишь в результате долгой эволюции стал скелетом. А сначала утолщение и упрочнение покровов у обитавших в морях далеких червеобразных предков членистоногих служило, надо полагать, приспособлением к защите внутренних органов от повреждений. Утолщение и уплотнение кутикулы делает ее менее проницаемой. Поэтому понятно, что только очень мелкие членистоногие, имеющие тонкие покровы, живущие в воде или в очень влажных местах, могут дышать всей поверхностью тела.
У всех более крупных членистоногих, имеющих и более плотные покровы, обязательно развиваются специальные дыхательные органы, поверхность которых покрыта такой тонкой кутикулой, что через нее легко может поступать кислород.
У живущих в воде членистоногих это жабр ы. Для дыхания в воде важно, чтобы поверхность жабр была возможно большей. Поэтому жабры
 | |||||
 | |||||
 | |||||
всегда представляют собой тонкие листовидные выросты или пучки пальцевидных отростков. Обычно жабрами у членистоногих становятся конечности или их части. В воде благодаря большой ее плотности даже тоненькие жаберные листочки не слипаются друг с другом. Поэтому на небольшом участке тела (в остальных местах защищенного плотными покровами) может разместиться много листовидных жабр.
Для доставки кислорода от жабр ко всем удаленным от них органам служит кровеносная система и полостная жидкость. У членистоногих кровеносная система представлена расположенным в спинной части тела пульсирующим, обычно продолговатым сосудом — сердцем. Кровь поступает в сердце через закрывающиеся клапанами боковые отверстия, а выталкивается из него в полость тела или непосредственно, или через немногие крупные сосуды — система кровообращения незамкнутая. У крупных морских членистоногих, имеющих толстый панцирь — у омаров, мечехвостов,— в крови есть дыхательный пигмент гемоцианин, сходный по химическому составу с нашим гемоглобином, но придающий крови не красный, а синеватый цвет: в состав гемоцианина входит не железо, как в гемоглобине, а медь. Гемоцианин легко присоединяет кислород при прохождении крови через жабры и отдает его внутренним органам. Так как у членистоногих кровеносная система незамкнутая, правильнее называть их «кровь» «гемолимфой», но и термином «кровь» зоологи широко пользуются.
На открытой поверхности суши ни кожное дыхание, ни дыхание с помощью жабр невозможно — легкопроницаемые участки покровов пропускают воду и быстро подсыхают, теряя проницаемость. Поэтому водные организмы, извлеченные на сушу, погибают от потери воды или от недостатка поступающего кислорода. На суше только скрытоживущие мелкие членистоногие, обитающие в таких местах, где воздух всегда насыщен водяным паром, например в почве, могут дышать всей поверхностью тела (мелкие клещи, ногохвостки, симфилы). Жизнь на суше вне влажных укрытий требует развития непроницаемых для испарения покровов, а такие покровы не пропускают и кислород.
У наземных членистоногих развиваются особые дыхательные органы — либо «легкие», представляющие глубокие мешковидные впячивания со складчатыми стенками, открывающиеся наружу лишь небольшими отверстиями (у пауков, скорпионов), либо (у большинства наземных членистоногих) трахеи. Трахеи представляют собой тонкие, обычно обильно ветвящиеся трубочки, открывающиеся наружу маленьким входным отверстием («дыхальцем») на поверхности тела. Ветви этих трубочек омываются полостной жидкостью, а самые мелкие ответвления доходят до отдельных
клеток. Из тонких трубочек трахей вода чер хальца испаряется очень медленно, и noil незначительны, а кислород по ним npd быстро и легко.
Уже далекие предки разных групп членУ гих приспособились к жизни в разных I Можно назвать группу водных членистя к которым относятся ракообразные, выдел] в подтип жабродышащих. Правда, среди сов| ных ракообразных есть и такие, которые жи суше (мокрицы), но по всем признакам онГ близки к своим водным родичам. Другой пои трахейнодышащие — представлен типично 1 ными членистоногими — различными груц многоножек и насекомых. Они прекрасно способлены к жизни на суше. В их покров! держатся воско- и жироподобные вещества,! щающие организм от высыхания, а все си органов, связанных с расходованием водщ делительная и дыхательная — действуют что влага расходуется минимально. Среди комых есть отдельные виды, проводящий жизнь или часть ее вводе, но строение таки ных насекомых явно показывает их происх ние от обитавших на суше предков.
За длившуюся сотни миллионов лет исто! группы обособились в связи с прпспособла к жизни в двух разных средах —в воде и на
Третий подтип членистоногих — хелицерЛ очень существенно отличается и от ракообра; и от трахейнодышащих. Среди хелпцеровыя и водные представители — мечехвосты, обита] в морях, и наземные — паукообразные.
В ископаемом состоянии сохранилось остатков хелицеровых. Их изучение, щее изучение современных представителей! зывает, как шло разделение классов в резу^ приспособления к жизни в разных средах -де и на суше. Изучение всех групп членист! убеждает в том, что переход их к жизни наГ был связан со способностью к выработке | храняющих от испарения влаги покровов, хранению в теле воды, без которой невоз5| жизнь.
У членистоногих, хорошо приспособив! к жизни на суше, панцирь бывает нередкГ ше, чем у обитающих в воде, но зато на его п| ности отлагается очень тонкий слой воскол ных и жироподобных веществ, не пропуска пары воды. Этот слой защищает организм а сыхания и препятствует испарению. Поэто всех членистоногих, живущих открыто наш ности суши, есть специальные органы дых! Те членистоногие, у которых специальных! нов дыхания нет, которые дышат всей по ностью кожи, вынуждены обитать во влажны рытиях — в почве, в лесной подстилке и только иногда по ночам и после сильных д| показываясь на поверхности.


 IУ наилучшим образом приспособившихся к шни на суше членистоногих чаще всего развива-■я трахейная система. Не только дыхательная, ридругие системы органов меняются у членисто-■их с переходом к жизни на суше. Выведение растваренных вредных продуктов крушения белков у водных членистоногих про-родит непосредственно во внешнюю среду через иверстия выделительных органов, открывающие-[я у основания конечностей головного или груд-отдела. А у всех членистоногих, наилучшим 1азом приспособившихся к жизни на суше, — у, многоножек, насекомых — выделительные 1убочки («мальпигиевы сосуды») открываются Iначале задней кишки. По мере прохождения по ей непереваренных остатков пищи и продуктов вделения из тех и из других стенки задней кишки йсывают воду, столь труднодоступную для жирей суши, а экскременты и продукты обмена родятся из кишечника в обезвоженном состоя-ш,
IУ наилучшим образом приспособившихся к шни на суше членистоногих чаще всего развива-■я трахейная система. Не только дыхательная, ридругие системы органов меняются у членисто-■их с переходом к жизни на суше. Выведение растваренных вредных продуктов крушения белков у водных членистоногих про-родит непосредственно во внешнюю среду через иверстия выделительных органов, открывающие-[я у основания конечностей головного или груд-отдела. А у всех членистоногих, наилучшим 1азом приспособившихся к жизни на суше, — у, многоножек, насекомых — выделительные 1убочки («мальпигиевы сосуды») открываются Iначале задней кишки. По мере прохождения по ей непереваренных остатков пищи и продуктов вделения из тех и из других стенки задней кишки йсывают воду, столь труднодоступную для жирей суши, а экскременты и продукты обмена родятся из кишечника в обезвоженном состоя-ш,
Приспособившиеся к жизни на суше членисто-
огие выработали способность запасать много жи-
[в особом образовании — жировом теле. Глав-
юроль жирового тела состоит в том, чтобы в кри-
рские периоды, когда организм не может полу-
■ъ воду извне, снабжать его... водой!
(При окислении жиров образуется много воды
ак называемая «метаболическая вода»), которая
(идет на покрытие расхода влаги животным.
этом отношении насекомые сходны с верблю-
1»ш, у которых при длительных переходах через
■иыню используется вода, образующаяся при
■ислении запасов жира в горбу. У водных групп
иешстоногих (ракообразных, мечехвостов) жи-
■рвого тела не бывает.
■Исключительно многообразны у членистоногих мы питания и характер пищи, от которых зави-■АЬроение ротовых органов и кишечника. При «коде к жизни в воздушной среде большую Воль в завоевании членистоногими суши сыгра-■о приспособление к питанию высшими растения-la. Цветковые растения черпают влагу из почвы ■сами становятся источником воды для наземных ■вотных. Представители многих групп насе-■шх, некоторые клещи и совсем немногие мно-■вожки и мокрицы способны к питанию живыми Ванями высших растений. Многие наземные ■шистоногие питаются разлагающимися ткапя-
■ растений, а многие — хищники или паразиты.
■ все они, завоевывая сушу, следовали за выс-
иши растениями — не только создателями пое-
■мых питательных веществ, но и живыми резер-
■арами влаги.
■ Наружный панцирь членистоногих настолько
■тен, что в затвердевшем состоянии не растя-
■вается. Поэтому рост членистоногих сопровож-
■ется линьками. У растущего животного
■арый панцирь становится тесным, лопается,
обычно сперва на спине, и сбрасывается. И только в короткий период, пока новый панцирь, образовавшийся под ним, не затвердеет, животное может увеличиваться в размерах. Во время сбрасывания старых покровов, которые сходят не только со всей наружной поверхности тела, но и со всех конечностей, в том числе и с ротовых органов, а также с передней и задней кишки и со всех дыхательных органов, членистоногие бывают вялыми, малоподвижными, не питаются и стараются спрятаться в каких-либо укрытиях.
Рост у членистоногих не непрерывный процесс, а ступенчатый, размеры тела увеличиваются скачкообразно — сразу после линьки. У многих членистоногих во время линьки происходит и большее или меньшее изменение строения тела. Часто взрослое животное приспособлено к совсем иному образу жизни, чем молодое, и потому особи разных возрастов (возрастом у членистоногих называется период между линьками) бывают даже совсем непохожи друг на друга. В таком случае развитие сопровождается метаморфозом. Превращение гусеницы в куколку, а куколки в бабочку может служить общеизвестным примером.
Образ жизни разных членистоногих неодинаков. Многие из них отличаются очень сложным поведением, что всегда связано с высоким развитием органов чувств и центральной нервной системы.
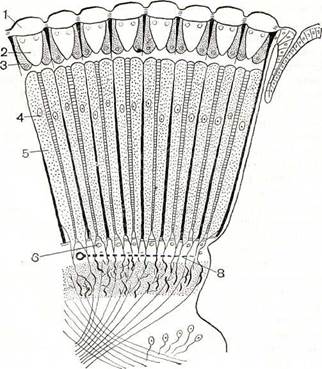
У большинства членистоногих хорошо развиты глаза. Глаза у членистоногих бывают простые, имеющие одну линзу, и сложные, или фасеточные. У высших представителей всех подтипов глаза сложные, фасеточные, состоящие из множества расположенных плотно рядом друг с другом маленьких глазков. Иногда число глазков (так называемых омматидиев) в фасеточном глазу измеряется несколькими тысячами (рис. 185). Фасеточные глаза характерны для высших ракообразных, мечехвостов и насекомых. Такие глаза были и у живших в палеозойскую эру трилобитов — членистоногих, полностью вымерших в конце этой эры. Каждый омматидий воспринимает только одну точку находящегося перед ним предмета (рис. 186). В результате сложный глаз дает изображение, состоящее из большого числа отдельных точек, т. е. мозаичное (как тоновое клише в книге) и прямое (рис. 187, см. т. 3, табл. 18). Вспомним, что изображение, полученное в нашем глазу, перевернутое и цельное!
Глаза членистоногих, оказывается, могут воспринимать не только те лучи, которые видимы для нашего глаза, но и ультрафиолетовую часть спектра. Например, многие цветы, которые нам кажутся равномерно белыми, для насекомых оказываются пестрыми; у таких цветов есть не видимый нами ультрафиолетовый рисунок (см. т. 3, табл. 19).


Рис. 185. Сложный глаз насекомого с вырезанным сектором (схема):
1 — линзы роговицы (фасетки); 2 — кристаллические конусы; 3 — главные пигментные клетки; 4 — ядра зрительных клеток; 5 — рабдомы.
Рис. 186. Схематический разрез через фасеточный глаз насекомого:
1 — линзы роговицы; 2 — кристаллические конусы; 3 — главные пигментные клетки; 4 — ядра зрительных клеток; 5 — рабдомы; 6 — нервные клетки сетчатки; 7 — рассматриваемый предмет; 8 — мозаичное изображение предмета в глазу.
И наземные и водные членистоногие способны различать обычный свет и так называемый поляризованный (те световые лучи, электромагнитные колебания которых совершаются в одной плоскости). Поскольку для рассеянного света неба характерна определенная степень поляризации лучей в меридиональной плоскости, членистоногие по небу, как по компасу, могут ориентироваться и в ясные и в пасмурные дни. Таким образом, членистоногие различают не только частоту, но и плоскость колебаний световых лучей.
Мы не ощущаем магнитные волны. А например, самки термитов в гнезде лежат так, что ось их тела направлена по магнитному меридиану, а если поместить рядом сильный магнит, меняют свое положение. Как воспринимают членистоногие магнитные волны, пока не выяснено. Отмечено, что членистоногие могут реагировать и на короткие радиоволны.
Хорошо развит и слух у членистоногих. Звуковые волны определенной длины (чаще короткие) они воспринимают с помощью тонких слуховых волосков на поверхности тела, особенно на усиках, с помощью специальных слуховых органов, напоминающих по строению струну, натянутую над резонаторной ямкой (как струна на балалайке). Часто наземные членистоногие издают ультразвуки, которых мы не слышим, с помощью различных «щщдуляционных» органов, по принципу действия напоминающих струну и смычок.
Особенно замечательна способность членистоногих воспринимать запахи — ничтожные концентрации многих веществ, растворенных в воде или взвешенных в воздухе. Никакие наши химические приборы не дают возможности уловить такие слабые концентрации вещества, а насекомые, например, могут по ним ориентироваться. Бабочки совки могут прилетать за километр на тарелку с бродящей патокой, а самцы шелкопрядов летя за километры на не ощущаемый нами и вблга запах коробки, в которой сидела самка. Не менее тонко восприятие и запахов, и содержания солей водными членистоногими. Многие наземные чле-1 нистоногие (мокрицы, личинки насекомых) способны улавливать разницу в относительной влаж-1 ности воздуха порядка 1—2%.
У водных членистоногих прекрасно развит органы равновесия.
Наиболее универсальное чувство, присущее всем членистоногим, — осязание. Осязательную функцию выполняют многочисленные волоски наI поверхности тела членистоногих. Для многих чле-1 нистоногих, особенно для донных, почвенных и нам вущих в тканях растений и животных, характер! стремление к наибольшей поверхности контакта t твердыми предметами — тигмотаксис.
Многообразная «информация», получаемая ш| внешнего мира органами чувств членистоноги, требует «обработки информации» центральной нерв-

|
У системой. Головной мозг — парные скопле-р нервных клеток над пищеводом — у высших итивных членистоногих очень хорошо развит, в р различается несколько отделов, связанных с ровными органами ориентировки в пространстве, расположенными--в головном отделе. Чем слож- т поведение членистоногого, чем многообразнее риовия, с которыми животное сталкивается, и чем ■ожнее ответы организма на воздействия среды, т более сложно строение головного мозга. Го-юной мозг у членистоногих связан окологло-§шым нервным кольцом с брюшной нервной це-■чкой. Исходным у членистоногих, по-видимому, ваяется соответствие каждому сегменту тела |рнганглиев брюшной нервной цепочки. Но при ■ияшш отдельных сегментов и даже отделов те-1 друг с другом происходит уменьшение чис-I нервных узлов, частично сливающихся друг ■другом, частично исчезающих. Степень расчле-1шя или слитности брюшной цепочки связана со шенью расчленения или слитности тела, и кон-■прация узлов брюшной нервной цепочки не ■кет служить показателем сложности поведения. IПоведение членистоногих, особенно в период ■няожения (спаривание, забота о потомстве), ■вает очень сложным, в чем можно убедиться из ■акомства с приводимыми ниже описаниями ■раза жизни разных представителей этого типа. иень часто поведение определяется прямой ■акцией на раздражение, например прибли-рие к источнику света (положительный фото-|иис) или уход от света (отрицательный фото-■сис). Еще более распространены реакции на ■здражение путем усиления или ослабления под-■жности (кинезисы). Например, многие насеко-I» активнее двигаются при более сильном или Вабом освещении и малоподвижны при среднем. ■им образом, после ненаправленного быстрого ■ижения они, наконец, попадают в благоприят-■е для себя условия освещения и перестают Ёиаться.
■роме непосредственных стандартных ответов 1 раздражения (безусловных рефлексов), для ■шистоногих очень характерны и сложные цепи ■следственно закрепленных рефлекторных дейст- ш (инстинкты). Инстинкты представляют такое ■южное поведение, какое в нормальных для дан-иго вида условиях является целесообразным, но ■о остается неизменным и при изменении усло-
■ и поэтому может становиться нецелесообраз-
ВНапример, роющие осы вырывают норку, при-■ш в нее парализованное насекомое, которым ■гана питаться личинка, откладывают на добы-
■ яйцо и заделывают норку. Все действия осы
■«сообразны». Но если в процессе деятельности
■в, когда она приступает к заделыванию норки,
■путь и добычу и яйцо, оса завершает свою уже
■ужную работу по замуровыванию норки.

Рис. 187. Изображение пламени свечи в глазу жука
светляка (Lampyris):
1 — в наружном слое фасеточного глаза, 2 — на сетчатке.
Наряду со сложными формами инстинктивного поведения у членистоногих (высшие ракообразные, пауки, высшие насекомые) прослеживаются и индивидуальные навыки; у них можно выработать и условные рефлексы. Мало того, например, пчелы могут передавать друг другу приобретенные навыки, обмениваться информацией. Определенными телодвижениями рабочая пчела сообщает другим пчелам в улье, в каком направлении надо лететь за взятком.
Членистоногим, несомненно, свойственны многие проявления высшей нервной деятельности, которую еще недавно считали характерной только для позвоночных.
Все членистоногие размножаются половым путем — обычно с оплодотворением, но иногда и без оплодотворения (партеногенетически). Большинство членистоногих раздельнополые, но некоторые, ведущие неподвижный образ жизни (например, усоногие ракообразные) или паразитические формы (например, некоторые равноногие ракообразные),— гермафродиты.
Типы оплодотворения у членистоногих разнообразны.
У водных членистоногих, например мечехвостов, бывает настоящее наружное оплодотворение, когда самка откладывает яйца в воду, а самец туда же выводит семенную жидкость. Проникновение сперматозоида в яйцо происходит при наружном оплодотворении во внешней среде — в воде.
Часто при наружном оплодотворении у водных членистоногих самец прикрепляет «пакеты» с семенной жидкостью (сперматофоры) к выводным половым отверстиям самок — откладываемые яйца оплодотворяются во внешней среде, но вероятность оплодотворения каждого яйца в таком случае увеличивается.
С переходом к жизни на суше низшие членистоногие, обитающие во влажной среде — в почве, в гнилой древесине и т. п. (многие клещи, скорпионы, ложноскорпионы, многоножки, ногохвостки и др.), выработали новый способ оплодотворения. Самцы откладывают сперматофоры во внешнюю среду, как при наружном оплодотворении (на
 землю, на остатки растений), а самки подбирают их своими половыми отверстиями, и, таким образом, проникновение сперматозоидов в яйцо осуществляется внутри организма самки, как при внутреннем оплодотворении. Такое оплодотворение называется наружно-внутренним.
землю, на остатки растений), а самки подбирают их своими половыми отверстиями, и, таким образом, проникновение сперматозоидов в яйцо осуществляется внутри организма самки, как при внутреннем оплодотворении. Такое оплодотворение называется наружно-внутренним.
У обитающих на поверхности суши в открытой атмосфере членистоногих оплодотворение внутреннее: самцы вводят семенную жидкость непосредственно в половые отверстия самок. Проникновение сперматозоида в яйцо происходит в защищенных от высыхания условиях внутри материнского организма.
В отдельных группах водных членистоногих, например у усоногих раков, также выработалась способность к внутреннему оплодотворению, повышающая плодовитость.
У большинства членистоногих наблюдается откладка яиц, живорождение встречается редко, например у некоторых двукрылых насекомых. Чаще наблюдается яйцеживорождение: самка рождает живых детенышей, однако развитие детенышей происходит за счет питательных запасов яйца, но не во внешней среде, а в яйцеводах самок (например, у тлей).
Как упоминалось, развитие многих членистоногих происходит со сложным превращением. В тех случаях, когда взрослая особь и форма, вышедшая из яйца, резко различаются по строению, молодые особи называются личинками.
У малоподвижных или неподвижных морских ракообразных расселяются подвижные личинки. У очень подвижных во взрослом состоянии крылатых насекомых личинки — в основном питающаяся стадия.
В тех случаях, когда яйца богаты желтком, из них выходят молодые членистоногие, уже похожие на взрослых. Такое развитие характерно для многих пресноводных ракообразных, паукообразных и некоторых насекомых.
Членистоногие ■— древние животные.
Самые достоверные данные, на основании которых можно судить о происхождении тех или иных групп животных, дают исследования ископаемых окаменевших остатков организмов или их отпечатков в слежавшихся и окаменевших наслоениях ила, песка или глины, под наносами которых трупы животных были погребены. Но не от всех прошедших эпох, когда уже существовала на Земле жизнь, сохранились ископаемые остатки организмов. Отложения самых древних периодов развития жизни на Земле подвергались в течение последующих эпох таким сильным изменениям, что точно установить, какие организмы обитали на Земле около миллиарда лет назад, невозможно, сколько-нибудь уцелевших остатков животных из отложений более древних, чем палеозойские, не сохранилось. А уже в самых глубоких («нижних») осадочных породах кембрийского периода
палеозойской эры (отлагавшихся более полуми лиарда лет назад) встречаются трилобиты (см. т. рис. 3) — животные с явно выраженными призн ками членистоногих, имевшие развитой наружнь скелет, расчлененные конечности, сложные гла; и другие свойственные представителям типа Arthri poda признаки организации. Трилобиты — по, тип членистоногих, полностью вымерший в кош палеозойской эры (длившейся 300—350 млн. лет
Из кембрийских отложений известны и ракоо! разные, некоторые из которых относятся к ccmpi менным отрядам, с силурийского периода извес ны родичи мечехвостов (см. т. 3, табл. 1) и да» скорпионов (см. т. 3, табл. 5, 1, 7), в девонски отложениях были найдены остатки ногохвоста а московский палеонтолог Б.Б.Родендор установил, что в девоне жили и крылатые насеш мые. Все эти данные свидетельствуют о том, чт тип членистоногих отделился от каких-то живо] ных, похожих на кольчецов, очень давно. Поэтом точно установить родственные связи различны членистоногих очень трудно. Несомненно, чт одной из первых оформившихся групп членисто ногих были трилобиты. В палеозое трилобиты до стигли расцвета, они буквально кишели в прибрея; ной зоне морей и океанов 500 млн. лет назад. Три лобиты сохранились в ископаемом состоянии на столько хорошо, что по некоторым остаткам уда лось не только исследовать их внутреннее стро ение, но на шлифах изучить даже микроскопическую структуру тканей этих животных, а такж(проследить их развитие. А применяя некоторьк методы современной химии, удалось даже выяснить, что в их крови был гемоцианин, как у современных высших ракообразных.
Из современных групп членистоногих наиболее близки к трилобитам, по-видимому, мечехвосты (см. т. 3, табл. 1), личинки которых несколько сходны с трилобитами, а зародыши — с личинками трилобитов («протаспис» — см. т. 3, рис. I
Мечехвосты и паукообразные имеют ряд общш черт строения, позволяющих их объединить в единый подтип хелицеровых (Chelicerata).
По характеру органов дыхания и ряду друга признаков некоторые общие с трилобитами черти имеют и ракообразные (класс Crustacea). Однако непосредственных родственных связей между трилобитами и ракообразными установить нельзя. Ракообразные представляют особый подтип жаб-родышащих (Branchiate) в типе членистоногих, А так как среди современных ракообразных мно-гие имеют более примитивное строение, чем хели-церовые, принято рассмотрение типа членистоногих начинать с них.
Последний подтип — трахейнодышащие (Тга-cheata) объединяет различных многоножек и насекомых, т. е. типично наземных членистоноги,]
В настоящее время не вызывает сомнений происхождение насекомых от каких-то древних членис-
 [еогих, близких к многоножкам, а многоножек— [очень примитивных червеобразных членисто-
[еогих, близких к многоножкам, а многоножек— [очень примитивных червеобразных членисто-
водные хелицеровые произошли от цих предков с трилобитами — это была другая вь эволюционного развития членистоногих, рспособившиеся к жизни в водной среде ра-
кообразные, сохранившие много черт древних членистоногих, представляют третью основнук ветвь развития членистоногих.
Знакомство с типом членистоногих и с его многообразными представителями удобнее всего начинать с подтипа жабродышащих, с класса ракообразных.
 ПОДТИП ЖАБРОДЫШАЩИЕ
ПОДТИП ЖАБРОДЫШАЩИЕ
 (BRANCHIATA)
(BRANCHIATA)
 ШООБРАЗНЫЕ______
ШООБРАЗНЫЕ______
pUSTACEA) '
[Бесконечно разнообразны водоемы, существую-ю на нашей Земле,— от необозримого океана, рина которого достигает почти 11 км, до малень-
| Е |
пересыхающих луж и капиллярных вод, продающих морской и речной песок. Различны температура, соленость, кислотность, газовый им п все остальные условия обитания в них ■х организмов.
'Немногие группы многоклеточных животных ■способились к жизни в этих столь разнооб-рых условиях. Большинство обитает или в мор-R,или в пресной воде, или на дне, или в толще р, ведет или свободный, или паразитический жизни. Этого никак нельзя сказать о рако-
■ЁЫХ.
Ракообразные населяют любые существующие 'Земле водоемы, пресноводные и морские, маккией большие, живут на дне и в толще воды, шикают в подземные воды, опускаются на превыше глубины Мирового океана. Большинство Ьообразпых свободно передвигается по дну н в толще воды, но среди них есть и сидяче-икреплеиные виды, например морские желуди юрские уточки, есть и паразиты, прикрепляются к рыбам и другим водным животным. Вод-I среда — царство ракообразных, однако не-иорые из них приспособились и к жизни на су-I Таковы общеизвестные мокрицы, некоторые юшческие крабы и раки-отшельники. шопе ракообразные чрезвычайно многочис-1ны. Морской планктон на всех широтах и на ■ глубинах в основном (не менее чем на 90% рассе) состоит из ракообразных. Очень велико
значение ракообразных и в планктоне пресныз вод. Донные ракообразные также нередко встречаются в массовом количестве.
Ракообразные играют очень важную роль в экономике природы. Органическое вещество в водоемах создается главным образом за счет жизнедея тельности микроскопических водорослей. Рако образные поедают эти водоросли и сами, в свок очередь, поедаются рыбами. Они выступают в ка честве посредников, делающих создаваемое в водо емах органическое вещество доступным для рыб С другой стороны, они используют в пищу огром пые массы погибших водных животных, обеспе чивая таким образом очищение водоемов.
Существование почти всех рыб, как морских так и пресноводных, в значительной степени зави сит от ракообразных. Некоторые рыбы, иаприме] сельдь, всю жизнь питаются планктонными рако образными, другие используют их только вскор(после выхода из икринки, а затем переходят ш какую-нибудь другую пищу. Для гигантов моря — беззубых китов — ракообразные служат основное пищей.
Многие ракообразные непосредственно исполь зуются человеком в качестве высокоценного пи щевого продукта. Во многих странах развит про мысел креветок, крабов, омаров, лангустов i некоторых других съедобных видов. Проведень успешные опыты по использованию морских планк тонных ракообразных для добывания витаминов жиров и других важных веществ. На рыбоводны: заводах разводятся некоторые виды ракообраз ных в качестве корма для молоди рыб.
Размеры тела ракообразных колеблются в очен: широких пределах — от долей миллиметра д< 80 см. Столь же разнообразны форма их тела строение его частей, окраска и другие приз наки.
В настоящее время известно около 30 000 ви дов ракообразных.
Тело ракообразных, так же как и других членистоногих, покрыто кутикулой, содержащей хитин. Она служит животным в качестве наружного скелета и защищает их от вредных внешних воздействий, но, с другой стороны, препятствует их беспрерывному росту. Поэтому время от времени ракообразные линяют — сбрасывают старую кутикулу и, пока не затвердела новая, быстро растут. Многие виды после линьки оказываются совершенно беспомощными и вынуждены укрываться от врагов в каких-нибудь убежищах.
Тело ракообразных состоит из отдельных сегментов, частично срастающихся друг с другом. Каждый сегмент снабжен парой членистых конечностей, но нередко в процессе эволюции некоторые конечности исчезают. Сегменты тела группируются в три отдела, имеющиеся и у других членистоногих,— голову, грудь и брюшко.
Голова представляет собой результат слияния головной лопасти и 4 следующих за ней сегментов и снабжена соответственно 5 парами придатков— передними и задними антенна-м и, находящимися у взрослых особей перед ротовым отверстием (только у Cephalocarida задние антенны расположены позади рта), жвалами, или мандибулами, и двумя парами ч е-л ю с т е й, или м а к с и л л. Обе пары антенн служат в первую очередь органами чувств. Они богаты различными чувствительными щетинками, осуществляющими функции обоняния, осязания и чувства химического состава воды. Жвалы размельчают пищу, челюсти фильтруют пищевые частицы и участвуют в процессе подачи пищи ко рту.
На голове помещаются также глаза. Обычно у ракообразных есть пара сложных фасеточных глаз, устроенных так же, как у других членистоногих. Иногда глаза сидят на стебельках,иногда непосредственно на поверхности головы. У некоторых ракообразных, например у ветвистоусых, сложные глаза сливаются в один непарный глаз. У личинки ракообразных — науплиуса — сложные глаза еще не развиты, но имеется один непарный простой, так называемый науплиальный глаз. У многих ракообразных этот глаз сохраняется и у взрослых, а у веслоногих он служит единственным светочувствительным органом в течение всей их жизни. У глубоководных и подземных видов глаза исчезают.
Задний край головы многих ракообразных снабжен покрытой кутикулой складкой, которая охватывает сверху и с боков весь грудной отдел тела или его переднюю часть. Это — кара-па к с, имеющий форму щита, двустворчатой раковины или полуцилиндра. В тех многочисленных случаях, когда карапакс отсутствует (жабро-ногие, веслоногие, бокоплавы, равноногие и др.), он исчез вторично. Карапакс не только служит
защитным образованием, но и определяет нап] ление токов воды, необходимых для питани дыхания.
Грудной и брюшной отделы тела у разных р; образных состоят из различного числа сегмен1 Нередко один или несколько передних груд сегментов срастаются с головой и их конечнс преобразуются в так называемые н о г о ч е i с т и, принимающие участие в захватывании пи
Брюшко заканчивается тельсоном, ответствующим пигидию многощетинковых < вей.
У наиболее примитивных ракообразных rj ные ножки выполняют три основные функцш двигательную, дыхательную и функцию поз пищи ко рту. Каждая ножка снабжена нескол! ми лопастями, расположенными на ее внутрен и наружном краях. Некоторые из наружных пастей превращены в жабры, другие лопасти зывают ток воды ко рту, третьи служат весла У более высокоорганизованных ракообраз] наблюдается разделение функций между разш конечностями: одни используются только передвижения, другие — только для дыхав третьи — только для захватывания пищи.
Чрезвычайно характерным для ракообраз] следует считать способ их дыхания. Как все стоящие водные животные, они дышат жабра через тонкие стенки которых растворенный в в кислород проникает в кровь. Даже сухопут: ракообразные — мокрицы имеют жабры и пользуют кислород смачивающего их тонкого с, влаги. Обыкновенно жабрами служат выро грудных ног, но у равноногих и ротоногих раке разных в жабры превращены брюшные конечное У части ракообразных (веслоногие, мистако риды, усоногие, ракушковые) жабр нет и дыха: совершается всей поверхностью тела.
Кровь приводится в движение биением серд которое помещается недалеко от жабр. У рано разных, жабры которых находятся на брюнп отделе, сердце помещается сзади, у остальных! расположено в грудном отделе. Кровеносные со ды есть только у представителей подкласса в ших ракообразных, у остальных кровь течет полостям между внутренними органами (р 188).
В крови часто растворены дыхательные и менты — красный гемоглобин и синий гемоц] нин, в котором железо заменено медью. Ды тельные пигменты могут связывать большое личество кислорода.
Питаются ракообразные различной пищей. (ни фильтруют имеющуюся в воде взвесь, включ! щую и мелкие организмы — бактерии, однок; точные водоросли и т. д., другие своими жвала откусывают куски мертвых и живых животе или растений. Пища перетирается жвалами ш падает в рот, а оттуда в пищевод. Задняя ча(
 рцевода у большинства ракообразных преоб-|ована в желудок, в котором иногда имеются [тикулярные зубы, завершающие измельчение щи. Экскременты удаляются через анальное [верстие, расположенное на тельсоне. Органами выделения ^служат железы, открываю-Ься наружу или при основании задних мак-рл(максиллярные железы), или щ основании задних антенн (антенная ь-не железы). Эти железы соответствуют [ганефридиям кольчатых червей. Выделяемая |и моча у многих пресноводных ракообраз-jx очень бедна солями. Животные удаляют из ■ости тела воду, и концентрация солей в их постной жидкости оказывается выше, чем в Ьужающей среде.
рцевода у большинства ракообразных преоб-|ована в желудок, в котором иногда имеются [тикулярные зубы, завершающие измельчение щи. Экскременты удаляются через анальное [верстие, расположенное на тельсоне. Органами выделения ^служат железы, открываю-Ься наружу или при основании задних мак-рл(максиллярные железы), или щ основании задних антенн (антенная ь-не железы). Эти железы соответствуют [ганефридиям кольчатых червей. Выделяемая |и моча у многих пресноводных ракообраз-jx очень бедна солями. Животные удаляют из ■ости тела воду, и концентрация солей в их постной жидкости оказывается выше, чем в Ьужающей среде.
1очти все ракообразные раздельной о-I только усоногие и некоторые десятиногие — рмафродиты. Обыкновенно самцы резко шчаются от самок внешним видом или размера-У одних ракообразных самцы крупнее, у дру- I мельче самок. Часто у самцов есть какие-ни-к> приспособления, помогающие им удержи-гь самку во время спаривания. Спаривание за-рчается в том, что самец прикрепляет близ (ювых отверстий самки сперматофор, I. кучку сперматозоидов, одетую оболочкой. |да самка начинает откладывать яйца, вместе ни выделяется вещество, растворяющее обо-|у сперматофора; сперматозоиды оказываются [свободе и оплодотворяют яйца. В других слу-Ь сперма вводится при помощи специальных улятивных органов непосредственно в половые самки. Яйца представителей подкласса жаб-югих ракообразных способны развиваться ртеногенетически, т. е. без опло-юрения. Имеются виды, состоящие из одних (ок, дающих начало новому поколению самок, рвнительно редко яйца откладываются прямо Ьу. Обычно самка носит их на себе в специаль-выводковой камере или прикреп-их к поверхности тела или к конечностям. |з яйца развивается характерная для всех ра-■азных личинка науплиус. Тело науп-Sa внешне не сегментировано. Он имеет три 1 конечностей — передние и задние антенны рндибулы. У некоторых ракообразных науп-k выходит из яйца и приступает к самостоя-юму суш,ествованию, у других эта стадия:руется внутри яйцевой оболочки и личинка дает яйцо, достигнув более поздней стадии штия. Наконец, есть ракообразные с прямым нтием, у которых все личиночные стадии про-рв яйце, а из него рождается уже сформиро-:ийся маленький рачок.
чинки ракообразных часто непохожи на лых и нередко ведут совсем иной образ жиз-Например, личинки многих донных ракооб-плавают в толще воды.
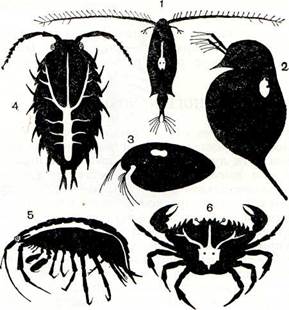
Рис. 188. Кровеносная система разных ракообразных:
1 — веслоногих, 2 — ветвистоусых, 3 — ракушковых, 4 — равноногих, 5 — бокоплавов, 6 — десятиногих.
Сравнение между собой по многим признакам различных ракообразных позволяет разделить этот огромный класс на 6 подклассов, к рассмотрению которых мы и переходим.
ПОДКЛАСС ЖАБРОНОГИЕ РАКООБРАЗНЫЕ
(BRANCHIOPODA)
Жаброногие ракообразные считаются самыми примитивными представителями всего класса ракообразных. Их голова не срастается с грудными сегментами. Грудные ножки листовидные, слабо хитинизированыые и служат одновременно для передвижения, дыхания и направления пищи ко рту. При их взмахах создается только один ток воды, который используется и для движения, и для питания, и для дыхания.
Число грудных сегментов непостоянно, строение половой, нервной и кровеносной систем примитивнее, чем у других ракообразных.
Жаброногие обитают преимущественно в пресных водах, морские виды немногочисленны и имеют пресноводное происхождение.
Все жаброногие ведут свободный образ жизни, среди них нет ни паразитов, ни сидяче-прикрепленных видов. Они не выработали приспособле-
ний к существованию в подземных водах и на больших океанических глубинах.
В этот подкласс объединяются два отряда ныне живущих ракообразных — жаброноги (Anostra-са) и листонозие ракообразные (Phyllopoda).








