 2015-08-12
2015-08-12 50819
50819Автор: научный сотрудник, Т. Морган, Научный сотрудник. Тип материала: Научная статья

Если допустить, что в одной хромосоме находится более одного гена, то возникает вопрос - могут ли аллели одного гена в гомологичной паре хромосом меняться местами, перемещаясь из одной гомологичной хромосомы в другую. Если бы такой процесс был невозможен, то гены комбинировались бы только в результате случайного расхождения негомологичных хромосом в мейозе. При этом гены, находящиеся в одной паре гомологичных хромосом, наследовались бы всегда сцепленно - одной группой.
Исследования Т. Моргана и его сотрудников, выполненные в начале XX века, показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером. В результате кроссинговера в гомологичных хромосомах возникают новые сочетания генов. Кроссинговер был обнаружен у всех организмов - животных, растений и микроорганизмов. Обмен идентичными участками между гомологичными хромосомами обеспечивает рекомбинацию генов. Это имеет большое значение для эволюции.
Кроссинговер можно обнаружить, учитывая частоты возникновения организмов с новым сочетанием признаков. Такие организмы называются рекомбинантами.
Явление кроссинговера было открыто на дрозофиле. Рассмотрим один из классических опытов Т.Моргана, позволивший ему доказать, что гены находятся в хромосомах в определенном порядке. У дрозофилы рецессивный ген черной окраски тела обозначают символом b, а его доминантный аллель, определяющий дикую серую окраску, - b+. Мутантный ген рудиментарных крыльев дрозофилы обозначают символами vg, а его нормальный аллель vg+.
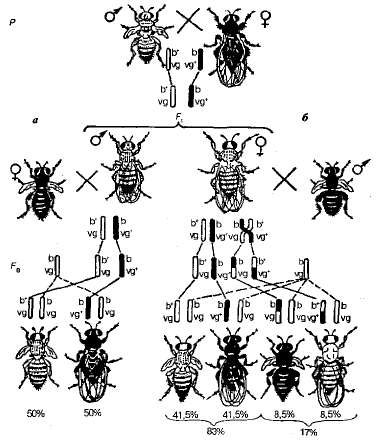
При скрещивании мух, различающихся по двум парам сцепленных признаков, - серых с рудиментарными крыльями и черных с нормальными крыльями - гибриды F1 по фенотипу будут серыми и с нормальными крыльями:

Полученных в первом поколении мух (отдельно - самцов и отдельно самок) Т. Морган скрещивал с мухами гомозиготными по мутантным аллелям - чёрными с рудиментарными крыльями.
Если гомозиготными по обоим рецессивным генам брали самок, а гибридными дигетерозиготами были самцы, то в потомстве получали расщепление в отношении 1 (серые с рудиментарными крыльями): 1 (черные с нормальными крыльями).
Такое расщепление показывает, что данная дигетерозигота обраpует гаметы только двух типов b+vg и b vg+. Сочетание генов в гаметах самца остаётся таким же, каким оно было у его родителей. Полученное расщепление показывает, что у самца не происходит обмена участками гомологичных хромосом. В дальнейшем выяснилось, что у самцов дрозофилы действительно, как в аутосомах, так и в половых хромосомах в норме кроссинговер не происходит. Поэтому при анализирующем скрещивании в потомстве появляются только две исходные родительские комбинации признаков в равных количествах. В данном случае наблюдается полное сцепление генов, находящихся в одной паре гомологичных хромосом.
Если для анализа взять гетерозиготными не самцов, а самок, то в Ра происходит другое расщепление. Кроме родительских комбинаций признаков появляются 2 новых типа – мухи с черным телом и рудиментарными крыльями, а также мухи с серым телом и нормальными крыльями.

В этом скрещивании сцепление тех же генов нарушается за счет того, что гены в гомологичных хромосомах поменялись местами благодаря кроссинговеру. Описанное явление называют неполным сцеплением генов.
Анализирующие скрещивания, описанные выше:
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными гаметами. Гаметы с хромосомами, не претерпевшими кроссинговер, называют некроссоверными гаметами. Соответственно организмы, возникшие от сочетания кроссоверных гамет гибрида с гаметами анализатора, называют кроссоверами или рекомбинантами. Организмы, возникшие от сочетания некроссоверных гамет с гаметами анализатора называют некроссоверными или нерекомбинантными.
При анализе расщепления по фенотипам было установлено, что в случае кроссинговера между двумя определёнными генами количественное отношение кроссоверных и некроссоверных классов всегда одно и то же. Обе исходные родительские комбинации признаков (серые с рудиментарными крыльями [b+ vg] и чёрные с нормальными крыльями [b vg+]), образовавшиеся из некроссоверных гамет, оказывались в потомстве анализирующего скрещивания в равном количественном отношении - примерно по 41,5%. В сумме некроссоверные мухи составили 83% от общего числа потомков. Два кроссоверных класса (чёрные мухи с зачаточными крыльями и мухи серые с нормальными крыльями) по числу особей были также одинаковы (по 8,5%). Суммарное количество кроссоверных особей было равно 17%.
Эти процентные соотношения сохранялись (с небольшими отклонениями в ту или иную сторону, обусловленными случайными причинами) при многократных повторениях этого эксперимента.
Законом сцепления Т. Моргана: гены, расположенные в одной хромосоме образуют одну группу сцепления и наследуются вместе.
Частота кроссинговера - это отношение числа кроссоверных особей к общему числу особей в потомстве от анализирующего скрещивания, выраженное в процентах.
Один процент перекреста был принят за единицу измерения частоты кроссинговера. Эта единица измерения в честь Т. Моргана была названа морганидой. Начиная с 80-х годов и в русскоязычной, и англоязычной литературе стали использовать термин сантиморган (краткое обозначение сМ). Таким образом, 1% кроссоверных особей = 1% перекрестов хромосом = 1% кроссинговера = 1 сантиморгану.
Величина перекреста хромосом отражает силу сцепления генов в хромосоме: чем она больше, тем меньше сила сцепления.
ЛИНЕЙНОЕ РАСПОЛОЖЕНИЕ ГЕНОВ В ХРОМОСОМЕ
Альфред Генри Стёртевант, ученик и сотрудник Т. Моргана, предположил, что частота кроссинговера отражает относительное расстояние между генами. Тогда, чем чаще происходит кроссинговер, тем дальше друг от друга расположены гены в хромосоме. Чем реже происходит кроссинговер, тем ближе друг к другу находятся гены.
Полагая, что частота кроссинговера зависит от расстояния между генами, А. Стёртевант проанализировал результаты многих экспериментов и в 1911 году установил ещё один закон наследственности - закон аддитивности. А. Стёртевант изучал частоты кроссинговера между тремя генами, расположенными в одной хромосоме (обозначим их А, В и С). Он установил, что если сравнивать частоты перекреста между генами А и В, В и С, А и С, то частота кроссинговера между двумя любыми из них, например А и С, близка к сумме его величин между генами А–В и С–В, т.е. АС% = АВ% + ВС%. Таким образом, наблюдается сложение расстояний между генами, определенных по частоте кроссинговера между ними. Эта закономерность соответствует обычной геометрической закономерности в расстояниях между точками на прямой. Отсюда, как следствие закона аддитивности, вытекало, что гены расположены в хромосомах в линейной последовательности и находятся на определенных расстояниях друг от друга.
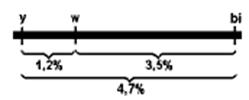
Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y, белый цвет глаз w и вильчатые крылья bi, были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w; 3,5% − от кроссинговера между генами w и bi и 4,7% — между у и bi.
Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно:
Многочисленные повторения описанных выше экспериментов Т.Моргана, выполненные другими генетиками, постоянно давали практически одни и те же результаты. Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т.е. каждый ген занимает в хромосоме свое определенное место. Фиксированное место, где расположен конкретный ген, получило название локус.
ОДИНАРНЫЙ И МНОЖЕСТВЕННЫЙ ПЕРЕКРЕСТЫ ХРОМОСОМ
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке и 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган предположил, что перекресты между двумя гомологичными хромосомами могут происходить одновременно в нескольких точках. Это предположение было им доказано на дрозофиле. Позже оно было подтверждено в экспериментах на других животных, а также растениях и микроорганизмах.
Кроссинговер, происходящий лишь в одном месте, называют одинарным, в двух точках одновременно - двойным, в трех - тройным и т. д., т.е. кроссинговер может быть множественным.
Пример. Были скрещены особи ABC/abc x abc/abc. Перекрест происходит вследующих точках между генами A и B, а также B и C.
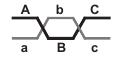
Результаты скрещевания:

Общая численность особей, полученных в этом опыте, — 521. Определяем число особей с одиночным перекрестом в участке 1: 37+42=79. К числу особей с одиночным перекрестом добавляем число особей с двойным перекрестом. Общее число особей с перекрестом в участке 1 равно 79+14=93. Выраженное в процентах от общего числа особей (521), это число отражает расстояние между локусами аллельных пар А—а и В—b, а также частоту перекреста. Таким же путем можно определить общее число особей с перекрестом в участке 2 (70+65+8+6=149). Следовательно, частота перекреста в участке 2 будет равна 28,60%. Необходимо принять во внимание, что 14 особей с двойным перекрестом учтены при подсчете частоты перекреста как в участке 1, так и в участке 2.
Другое обстоятельство, которое следует иметь в виду в отношении двойных перекрестов, состоит в том, что они затрагивают только среднюю часть хромосомы между локусами А—а и С—с. Таким образом, при двойных перекрестах изменяется только положение генов В и b, а расположение локусов А—а и С—с остается неизменным. Если мы не проконтролируем наследования генов В—b, то будет невозможно определить наличие двойных перекрестов. Частота перекрестов, определяемая непосредственно по сцеплению между генами А и С, без учета передачи генов В—b будет менее достоверной. В нашем примере всего у 214 особей из 521 обнаруживается перекрест между локусами А и С, следовательно, частота его равна 41,07%. Эту величину можно сравнить с суммой вычисленных ранее значений для перекрестов в участках 1 и 2. Эти значения были равны 17,85 и 28,60%, что дает в сумме 46,45%, т. е. на 5,38 единицы больше величины, полученной непосредственным определением частоты перекреста между локусами А и С.
Определение расстояния от А до С осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов (41,1%) прибавляют удвоенный процент двойных кроссоверов (2,7x2 = 5,4%). Удвоение процента двойных кроссоверов необходимо потому, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках. Поэтому, чтобы вычислить процент одинарного кроссинговера, необходимо величину двойного кроссинговера умножить на 2.
ИНТЕРФЕРЕНЦИЯ
Интерференция - это явление, при котором кроссинговер, произошедший в одном участке хромосомы, препятствует перекресту хроматид в близлежащих участках конъюгировавших хромосом. Установлено, что в опыте процент двойных кроссоверных особей часто оказывается ниже теоретически ожидаемого. Одной из причин, снижающих наблюдаемую величину кроссинговера, оказывается процесс подавления второго кроссинговера вблизи того пункта, где обмен уже произошел. Кроссинговер, произошедший в одном месте хромосомы, подавляет кроссинговер в близлежащих районах. Это явление носит название интерференции. Особенно сильно сказывается интерференция на подавлении двойного кроссинговера при малых расстояниях между генами. Если гены А, В и С близко расположены близко друг к другу, то одинарный обмен на участке между генами А и В подавляет кроссинговер на участке между В и С. Разрывы хромосом оказываются зависимыми друг от друга. Степень этой зависимости определяется расстоянием между происходящими разрывами: по мере удаления от места разрыва возможность другого разрыва увеличивается.
Величина интерференции может быть измерена. Для этого необходимо маркировать хромосому на большом протяжении генами, место и последовательность расположения которых известны. Зная место и последовательность расположения генов в хромосоме, можно рассчитать теоретически ожидаемую частоту двойных перекрестов. Величина интерференции измеряется отношением числа наблюдаемых двойных разрывов к числу возможных двойных разрывов при допущении полной независимости каждого из них.
Объясним это на рассмотренном ранее примере. Было установлено, что гены А и В разделены расстоянием 17,9 сМ, а В и С - расстоянием 28,6 сМ. Если разрывы на участках АВ и ВС происходят как независимые друг от друга и случайные события, то вероятность двойного кроссинговера между генами А и С должна быть равна произведению процентов кроссинговера на участках АВ (17,9%) и ВС (28,6%), т.е. (17,9: 100) х (28,6: 100) х 100% = 5,12%
Но в опыте мы получили среди 521 особи всего 14 особей, возникших как следствие двойного кроссинговера, что соответствует 2,68%. Полученный в опыте процент значительно ниже ожидаемого. Это снижение и объясняется наличием интерференции.
Итак, интерференцию измеряют отношением наблюдаемого числа двойных, перекрестов к теоретически ожидаемому. Это отношение называют величиной совпадения или коинциденцией, и выражают в долях единицы, или в процентах. В приведенном примере коинциденция равна 2,68:5,12=0,52, или 52%.

