 2015-06-28
2015-06-28 4989
4989Мейоздық айқас. Генетикалық әдістер арқылы хромосомалардың айқасу құбылысы ашылғанға дейін, цитологтар мейоздың профазасын зерттей отырып хромосомалардын, өзара біріне-бірі оралатынын, сөйтіп, % тәрізді фигура—хиазма түзетінін байқады. 1909 жылы Ф. Янсенс хиазмалар хромосомалар учаскелерінің алмасуымен байланысты деген болжау айтты. Кейіннен бұл көріністер 1911 ж. Т. Морган ұсынған хромосомалардың генетикалық айқасу жөніндегі гипотезаға қосымша негіз болды.
Хромосомалар айқасуының механизмі мейоздың I профазасындағы гомологиялық хромосомалардың күйімен байланысты болады. Мейоз үшінші тарауда жан-жақты қарастырылған. Оның ерекшеліктерін еске түсірейік. I профазада гомологиялық хромосомалар сәйкес учаскелерімен конъюгацияланады. Әр хромосома бивалент кезінде екі хроматидтен түрады, ал бива-ленттің езі тиісінше төрт хроматидтен тұрады (V таблица, А). Сонымен коньюгация — гомологиялық хромосомалар арасында кроссинговер жүре алатын жалғыз кезең. Сонымен кроссинговер төрт хроматид стадиясында өтеді жене ол хиазмалар түзілу кезіне сәйкес келеді.
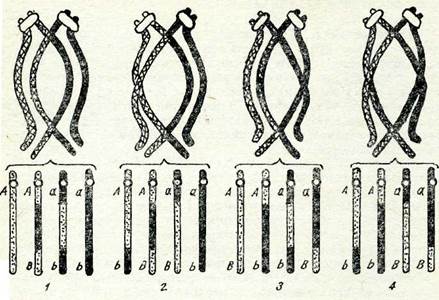
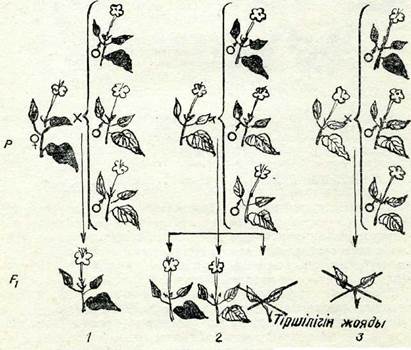
Егер бір бивалентте бір алмасу емес, екі және одан да кеп болса, онда бірнеше хиазмалар түзіледі. Бивалентте төрт хроматид болатындықтан, анығында олардың әрқайсысы басқа хроматидтердің кез келгенімен бірдей мүмкіндікте учаскелерін алмастыра алады. Бүл кезде алмасуға екі, үш немесе тәрт хро-матид қатыса алады. 50-суретте осындай алмасудың схемасы берілген: 1) сіңлілі емес екі хроматид арасындағы реципрокты қос қабат алмасу, егер гендер — маркерлерде алмасу болмаса, бүлар гендер рекомбинацияларын бермейді (50-сурет, 1); 2) диагональды алмасу, бүл жерде әр түрлі екі ауданда сіңлілі екі хроматид сіңлілі емес белгілі бір хроматидпен бір мезгілде бір қабат айқасуға түседі, ал төртінші хроматид алмасуға кіріспей-ді. Осындай қос кабат алмасу нәтижесінде үш рекомбинантты хромосома пайда болады, ал біреуі рекомбинатты емес. күйінде қалады. (50-сурет, 2, 3); 3) комплементарлы алмасу бүл, кезде терт хроматидтің тертеуі де әр түрлі ауданда бір қабат алма-сады, мүнда төрт хроматидтің сіңлілі емес екі хроматиді жұп-тасып бір орында бір қабаттан алмасады, ал басқа екеуі басқа учаскеде бір қабаттан алмасады, осының нәтижесінде рекомбинантты терт хромосома түзіледі (50-сурет, 4). Бұнда алмасуға үш хроматид қатысса, хроматидтер арасында бір мезгілде бір қабаттан алмасу болса, қос қабат кроссоверлер пайда болады.
 |
Осы уақытқа дейін сіңлілі емес хроматидтер арасындағы кроссииговер қарастырылды, сіңлілі хроматидтер ішіндегі алма-су рекомбинациялар бермейді, осыған байланысты комбинативтік өзгергіштіктің биологиялық механизмі ретінде мүндай алмасудың маңызы жоқ.
50-сурет. Қос қабат хиазм схемасы мен хроматидтер арасындағы кос қабат алмасу өнімі:
1 — сіңлілі емес хроматидтер арасындағы реципрокты ңос ңабат алмасу (екі жіп-ше алмасқан). 4 —барлык, хроматидтер арасындағы комплементарлың алмасу (төрт тікше алмасңан); 2, 3 —үш хроматид арасындағы диагональды алмасу (үш шіпше алмасқан).
Соматикалық (митоздық) кроссинговер. Кроссинговер мей-оздың I профазасында гамета түзілу кезінде болатынын біз жоғарыда айттық. Бірақ соматикалық немесе митоздық крос-синговер деген де бар, олар дене клеткаларының митоздық бө-лінуі кезінде, көбінесе эмбрион тканьдарында болады.
Митоздың профазасы кезінде гомологиялық хромосомалар әдетте коньюгацияланбайды және бірінен-бірі тәуелсіз орна-ласады. Бірақ кейде гомологиялық хромосомалардың синапсисі және хиазмаға ұқсас фигураларды байқауға болады, бұл кез-де, әрине, хромосомалар санының редукциясы байқалмайды.
Соматикалық кроссинговер белгілердің көрінісін, ала-құла-лыққа (мозаичность) жеткізуі мүмкін. Ол құбылыстың меха-низмін мынадай мысалдан анық түсінуге болады. Тіркескен рецессивті гендері у (дененің сары бояуы) және зп (бүріскен қылшықтар) боиынша гетерозиготалы "■=—= дрозофила ұрға- ут8П шыларының денесі сүр және қылшықтары қалыптағыдай болады. Егер дене клеткаларындағы хромосомалардың осы жұбы айқасса, онда оның денесінде рецессивті белгілері бар дақтар пайда болады (51-сурет, 1).
Схемадан көрінгендей, (51-сурет, 2) егер соматикалық кроссинговер сіңлілі емес екі хроматидтер арасында жүрсе, оны та
| 51-сурет. Дрозофила денесінің алақұла (қос қабат дағы бар) учаскесі, ол құбылыс—денелік кроссинговердің нәтижесі: |
буға болады. Егер алмасу ориы зп гені мен центромера арасында болса және егер митоздық сол циклдің анафазасында екі хромосома узп+ кездейсоқ бір полюске, ал екі хромосома у+зп екінші полюске кетсе, онда соматикалық кроссинговер қос дақ түрінде болады. Бір митоздың нәтижесінде түзілген екі клеткадан көбею кезІнде
тканьдерде үлкендігі шамалас екі түрлі дақ пайда болады және олар рецессивті белгілер береді, өйткені у жәке зп гендері болып өткен
| 1 — шыбын торансіндегі дененің сары (ақыл) бояуы, қлыпты қылықары бар және «от шалғандай» дылшыңтары бар, біраң ол сұр түсті. 2 — соматикальщ кроссииговер схемасы: түзілген клеткалар генотиитерінің өзара айырмасы бар және бастапқы формадан да айырмасы бар. |
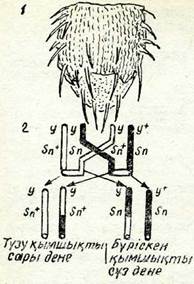 |
Шыбынның қалыпты қылшығы бар сүр түсті, денесінде сары түсті бір дақ, қылшықтары «бүріскен» сүр түсті екінші дақ пайда болады. Дақтардың пайда болуы тек төрт жіпше стадиясы кезіндегі алмасу-дың нәтижесі болғандықтан, бұл жағдай кеп клеткал
организмдерде кроссинговердің хроматидтер ара-сында жүретіндігінің тікелей дәлелі болып табылады.
Кроссинговер механизмі жөніндегі гипотезалар. Хроматидтердің
айқасуы механизмі женінде бірнеше гипотеза бар, бірақ бүлар-дың біреуі де гендердің рекомбинациялану фактілерін және бүл кезде байқалатын цитологиялық көріністі толық түсіндіре ал~ майды.
Ф. Янсен үсынған және К. Дарлингтон ары қарай дамытқан гипотезаға сәйкес, гомологиялық хромосомалар синапсисі процесіндегі бивалент кезіндегі хромосома жіптерінің спиральда-дануына, сол сияқты бивалент кезінде гомологтардың бірін-бірі орап айқасуына байланысты пайда болатын динамикалық кернеу түзіледі. Осындай кернеу нәтижесінде төрт хроматидтің біреуі үзіледі. Бүл үзілу биваленттегі тепе-теңділікті бүза оты-рып, осы биваленттегі басқа бір хроматидте дәл соыдай иүктеде теңестіруші үзіліске себеп болады. Бүдан кейін хроматидтердің үзілген ұтары реципрокты жалғасып кроссинговер болуға әке-леді. Осы гипотезаға сәйкес хиазмалар кроссинговермен тіке-лей байланысты.
К. Сакс гипотезасы бойынша хиазмалар кроссинговердің нә-тижесі емес: ең алдымен хиазмалар түзіледі, одан кейін алмасу.
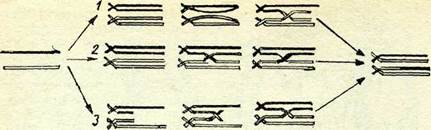
52-сурет. Хромосомалар айқасуы механизмнің әр түрлі гипотезаларын көрсе-туші схемалар:
1 — хромосомалардың үзілгеынен кейін ңайта комбинациялануы; 2 — хиазм түзілуі және одан кейін хромосоманың үзіліп алмасуы; з — көшірмесін таңдау кездегі қа-телер.
жүреді. Хромосомалар полюстерге тараған кезде механикалық кернеу салдарынан хиазмалар болған нүктелерде үзілістер және сәйкес учаскелерде алмасулар жүреді. Алмасудан кейін хиазма жойылады.
Д. Беллинг ұсынған және И. Ледерберг толықтырып жаңартқан басқа гипотезаның мағынасы мынадай: ДНК-ның репликациялану процесі бір жіпшеден басқасына реципрокты ауыса алады; бір матрицада басталған өзін өндіру процесі белгілі бір нүктеден гомолог партнер ДНК-ның матрица жіпшесіне ауыса-ды, бүдан кейін өзінің матрицасына қайтып оралады, бұл жағ-дай генетикалық материалдың рекомбинациясына алып келеді. Бүл гипотеза «көшірмесін таңдау» гипотезасы деп аталды.
Ақырында К. Уайтхауз бен Р. Холлидей кроссинговерді мо-лекулалық процесстер деңгейінде түсіндіруге әрекет жасады (17-тарауды қараңыз). Ол гипотезалар схема түрінде 52-сурет-те берілген. Д. Тейлор кроссинговердің үзілу механизмінің пай-дасына дәлелдер алды.
10. ХРОМОСОМАЛАРДЫҢ АЙҚАСУЫНА ӘСЕР ЕТЕТІН ФАКТОРЛАР
Кроссинговер процесіне эсер ету мумкіндігі. Хромосомалар клетканың бүкіл системасының ажырамас бөлігі болып табылады. Хромосомалардың редупликациясы және бөлінудің бүкіл циклі барлық клетка әрекетінің нәтижесі болып табылады.
Хромосомалардың айқасуы физиологиялық, физикалық және биохимиялық күрделі процесс ретінде, оған сыртқы ортаның факторлары, түтас организмнің және жеке клеткалардың қызмет ету күйі әсер етеді. Хромосомалардың айқасуын сол сияқты олардың ішкі қүрылымы мен организмнің, генотипі қамтамасыз етеді.
Гомо-және гетерогаметалы жыныстағы айқасу. Зерттелген жануарлардың және өсімдіктердің көпшілігінде хиазмалар- мен айқасу екі жыныста да мейоз кезінде орындалады. Бірақ мейоз-дық кроссинговер гомогаметалы жыныста ғана болып, ал гетерогаметалы жыныста қалыпты жағдайда ол болмайтын жану-арлардың жекелеген түрлері болады. Мұнын. гетерогаметалы жыныста синапсистің болмауын олардың гетероморфтылығымен түсіндіруге болатын жыныс хромосомаларына ғана емес, соны-мен бірге аутосомаларына да қатысы бар. Дрозофиланың ер-кектерінде және жібек құртының ұрғашыларында мейоздың I профазасында синапсис және хиазмалар пайда болмайды. Сол түрлердің гомогаметалы жынысында (дрозофиланың үрғашыла-рында және жібек құртының еркектерінде) хромосомалар ай-қасуы қалыпты өтеді.
Хромосомалар құрылымының айқасу жиілігіне әсері. Митоз-да және мейозда хромосомалардың біртұтас қүрылымды болу-ын центромералар қамтамасыз етеді. Әдетте центромералардың қасында гетерохроматинді учаскелер болады.
Хромосомалар айқасуында да центромера аса маңызды роль атқарады. Дрозофилада центромера маңында әдетте айқасу си- рек кездеседі. Центромерадан қашықтаған сайын айкасулар жиілейді, бүдан кейін үшына таман қайтадан азаяды. Сондықтан дрозофиланың генетикалық карталарында центромераға қашық аудандарға қарағанда оған жақын жерде гендердің шо ғырлануы көп болады. Жүгеріде кроссинговерге центромераның мүндай әсері табылмады.
Хромосоманың әр түрлі учаскелеріндегі айқасу жиілігіне гетерохроматинді және эухроматинді аудандардың бөлінуі әсер етеді. Гетерохроматинді учаскелерде сыртқы орта факторлары-ның әсерінен айқасу жиілігінің үлкен өзгеріске үшырайтыны байқалады. Кроссинговер үшін гетерохроматинді және эухрома-тинді аудандардың маңызы, осы аудандардағы хромосома жіптерінің спиральдану дәрежесімен байланысты болуы мүмкін. Центромера ауданында кроссинговер жиілігінің төмендеуі, осы аудандағы хромосомалардың жақсы спиральдануы салдарынан да болуы мүмкін. Спиральдану гендердің цитологиялық ара қашықтығын азайтады, ал оның күшеюі гомологиялық аудандардың синапсисіне және айқасуына кедергі жасай алады.
Организмнің қызмет ету жағдайымен генотипінің хромосо-малар айқасына әсері. Кроссинговердің жиілігі организмнің жасына тәуелді. Егер генетикалық картада өзара жақын орна-ласқан, мысалы бірінен-бірі 60% қашықтықта орналасқан екі геннің арасындағы айқасуды зерттесе, және кроссинговер про-центін дрозофила үрғашыларынан әр он күн сайын есептесе (1 — 10, 11—20, 21—30 күн бойынша), онда айқасу жиілігі тиісінше 5,9; 1,8жәнеЗ,8% болады.
Организмнің қызмет ету күйі мейоздың әр түрлі стадиясының барысына әсер етуі мүмкін (демек хро.мосомалардың айқасуына да), өйткені хромосомалардың спиральдану дәрежесі, профазаның әр түрлі стадиясының өту жылдамдығы клеткалардың физиологиялық күйіне өте тәуелді болады.
Кроссинговердің жиілігіне генотип әр түрлі жолмен әсер ете алады. Мысалы хромосомалардың қайта құрылуы олардың
айқасу жиілігіне қатты әсер етеді, өйткені ондай қайта құрылу зигонема стадиясында хромосома синапсисінің қалыпты барысын бұзады (13-тараудан қараңыз). Әр түрлі объектілерде мей-оздың I профазасындағы хромосомалар синапсисін, спиральда-нуын және мейоздағы хромосома редукциясын, хиазмалардың түзілуін бақылайтын гендер табылды. Анығында, осындай ген-дер го.мологиялық хромосомалардағы алмасу жиілігіне де әсер ете алады.
Сыртқы орта факторларының айқасуға әсері. Осы уақытқа дейін біз кроссинговердің генотипке және ол анықтайтын клетка мен организмнің физиологиялық күйіне тәуелді екенін қарастырдық. Айқасудың мүндай типін спонтанды кроссинговер деп атайды. Бірақ сыртқы ортаның әр түрлі факторларымен организмге әсер ету арқылы айқасу жиілігін өзгертуге болады.
Әр түрлі факторлармен қолдан әсер ету нәтижесінде болған айқасуды индукциялаған кроссинговер деп атайды. Хромосомалардың айқасуына көптеген факторлар әсер етеді: жоғары және төменгі температура, иондаушы сәулелер, клеткаларда кальций, магний т. б. иондардың болуы, мысалы дрозофилада төменгі (+9... + 13°С) және жоғары (+ 30... +32°С) темпера-тура кроссинговер процентіи көбейтеді; даму ең қолайлы тем-пературада өткенде айқасу проценті өте аз болады. Хромосомалардың центромераға қашық орналасқан бөліктерімен салыс-тырғанда оған жақын орналасқан аудандары сыртқы орта әсе-ріне тез үшырайды. Иондаушы радиация мен химиялык атенттер де кроссинговер жиілігін арттырады. Мысалы, этилендиаминтетрауксус қышкылы хромосомадан кальций және магний иондарын бөліп шығарады, сірә, ол иондар хромосомалардың құрылым тұтастығын сақтауға белгілі бір роль атқаратын болса керек, сондықтан оларды бөліп шығару хромосомалар құылымының тұастық күйін бұзады, бүл жағдай хроматидтердің үзілу жиілігін арттырады, ал ондай үзілген хроматидтердің бір бөлігі гендердің рекомбинациялануына алып келеді.
Сыртқы факторлардың кроссинговерге әсер етуінің нақты механизмі әлі анықталған жоқ.Жыныспен тіркесіп түқым қуалауды және кроссинговер құбылысын зерттеу, алғашқы күйінде Морган үсынған тұым қуалаудың хро.мосомалык теориясын тұжырымдауға мүмкіндік берді.
Жоғарыда айтылғанға мынаны қосуға болады: айқасудың нәтижесінде гендер комбинациялана алады және белгілердің үйлесімдері пайда болып ол үйлесімдер организмдердің ортаға жақсы бейімделуін қамтамасыз етеді. Сонымен, кроссинговер комбинативтік өзгергіштікті арттыра отырып эволюция проце-сінде және селекцияда аса маңызды роль атқарады.
10-тарау. ХРОМОСОМАСЫЗ (ЦИТОПЛАЗМАЛЫҚ) ТҰҚЫМ ҚУАЛАУ
Белгілі бір структура тұқым қуалаудың материалдық сақтаушысы қызметін атқара алуы үшін және тұқым қуалаудың сандық заңдылықтарын қамтамасыз ету үшін жоғарыда айтылғандай (3-тарауды қараңыз), оның негізгі үш қасиеті болуы керек: клетка метаболизмінде аса маңызды қызмет атқаруы тиіс, өзін-өзі өндіретін қабілеті болуы керек, беліну кезінде пайда болған жас клеткаларға теңдей дәл таралуы керек. Осы үш шартты ядроның құылымы — хромосома толық қанағаттандыра алады.
Цитоплазманың көптеген органоидтары бірінші шартты қа-нағаттандырады. Мысалы, клетка бөлінуі кезінде центриольдар ұшық түзуге қатысады, пластидтер аса маңызды синтездеу процесін қамтамасыз етеді, митохондрия оның тыныс орталығы болып табылады, рибосомаларда белок синтезделеді т. с. с. Центриольдар, пластидтер және митохондриялар өздерін жа-ңартып жасап шығара алады, яғни екінші шартты да қанағат-тандырады.
Бірақ цитоплазманың бірде-бір органоиды (центриольды қоспағанда) клетканың бәлінуі кезінде хромосомалар сияқты дәл таралмайды. Ядро қүрылымдарының (хромосомалардың) цитоплазма құрылымдарынан негізгі айырмасы осы қасиетінде.
Ядро мен цитоплазма арасында бүдан езге де айтарлықтай айырмашылық бар: 1) ядрода әр түрге тән және шектелген хро-мосомалар саны болады; цитоплазмада әдетте бір мағыналы көп органоидтар болады, олардың саны түрақты болмайды; 2) көпшілік жағдайда ядро хромосомаларда пайда болған же-тіспеушілікті түзетіп, орнына келтіре алмайды, ондайлар клет-ка бөліну кезінде орнына келеді; жетіспеушілікке үшыраған және өзі көбейе алмайтын цитоплазма органоидтарының орнын жетіспеушілікке ұшырмаған сондай органоидтар қалпына келтіре алады.
Хромосомалар мен цитоплазма органоидтары. қасиеттерінің арасындағы келтірілген айырмашылық, клеткалардың осы эле-менттерімен анықталатын тұқым қуалау заңдылығына негізде-луі керек. Хромосомамен анықталатын тұқым қуалау ядролыщ немесе хромосомалық деп аталды. Егер түқым қуалаудың ма-териалдық негізі цитоплазма элементтері болса, ондай түқым қуалау хромосомасыз немесе цитоплазмалыщ деп аталады.
Өсімдіктердің және жануарлардың жүмыртқа клеткасында цитопл.азма көп, ал аталық гаметада әдеттегідей цитоплазма жоққа тән болғандықтан, мынадай жағдайды күтуге болады: хромоеомалық түқым қуалаудан цитоплазмалық түқым қуалау өзгеше, аналық саламен іске асуы тиіс.
Екіншіден, клетка бөлінуі кезінде цитоплазма органоидтарын бөліп ажырататын хромосомадағыдай дәл механизм болма-ғандықтан,
цитоплазмалық тұқым куалауда, ядролық тұқым куалаудағыдай қатал сандық заңдылықтардың сақталуы жоқ.
1. ЯДРО МЕН ЦИТОПЛАЗМАНЫҢ ТҰҚЫМ ҚУАЛАУ КЕЗІНДЕГІ САЛЫСТЫРМАЛЫ РӨЛІ
Андрогенез кезіндегі тұқым қуалау. Тұқым қуалаудағы ядро-ның және цитоплазманың салыстырмалы ролін анықтауға мүм-кіндік беретін сенімді әдіс — цитоплазмасы бір түрге жататын,. ал ядросы екінші түрге жататын клеткалар алу.
Ядроны алмастырудың бір әдісі — бір түрдің жұыртқа клеткасындағы ядросын жойып, оны екінші түрдің сперматозоидымен ұрықтандыру. Мұндай гаплоидты зиготалар дамығанда гаплоидты андрогенді особьтар пайда болуы тиіс, өйткені ондайг особьтар аталық ядро мен аналық цитоплазмадан шыққан. Бірақ гаплоидты зиготалар әдетте эмбриональдық стадияның ер-терек кезеңінде өледі. Сондықтан Б. Л. Астауров жібек құты-ның екі ВотЬух тогі және В. тапсіагіпа түрлерін шағылыстыру арқылы диплоидты андрогенді будандар алу жолын іздеді.
Жібек көбелегінде гетерогаметалы жыныс (ХУ) ұрғашысы,, ал гомогаметалы жыныс (XX) еркегі. Онан басқа көптеген на-секомдардағы сияқты, жібек көбелегінде де полиспермия қүбы-лысы байқалады.
Жібек көбелектерін шағылыстыруда В. тапсіагіпаХВ. тоіі еркегінің әр түрлі хромосомаларының рецессивті үш гені таң-баланды, сһ — личинкаларының сарғыштау қоңыр түсі, ті — жұлдыз қүрттарының сүттей ақ түсі, р — көбелектердің ақ түсі. Үрғашысының тиісінше үш доминантты ген-личинкалардың қа~ ра реңін анықтайтын, жұлдыз қүрттардың сүр түсін, көбелектер-дің қоңыр түсін анықтайтын гендері болды (53-сурет). Демек, мұндай шағылыстыру кезінде қалыпты будан үрпақтардың бә-рінің личинкалары қара түсті, жүлдыз құрттары сүр түсті және көбелектері қоңыр түсті болуы керек, ал андрогенді особьтарда мынадай рецессивті белгілер пайда болуы тиіс: личинкалар сар-ғыш қоңыр түсті, жүлдыз құрттар мен көбелектер ашық түсті болуы керек.
Жүмыртқа клеткадағы ядроны бұзу екінші мейоздық бөліну кезінде 40°С температурамен әсер ету арқылы жүргізілді. Әр жолы салынған гренаның бір бөлігіне 40°С температурамен әсер етілді, екінші белігі бақылау үшін қалдырылды. ТәжірибедегІ топта аналық ядро елтірілгендіктен, екі аталық пронуклеус әза-ра құйылысып, диплоидты бір ядро түзген жағдайда ғана жаңа организмнің дамуы жүруі мүмкін. Осының нәтижесінде дамы-ған барлық особьтар еркек жынысты (XX) және олардың ре-цессив белгілері болды, өйткені еркек организм рецессивті ген-дері бойынша гомозиготалы болды (53-сурет). Осы тәжірибенің нәтижелерін түқым қуалауда ядроның жетекші роль атқаратын-дығының және цитоплазманың айтарлықтай әсері болмайтын-дығының тікелей дәлелі ретінде қарастыруға болады.
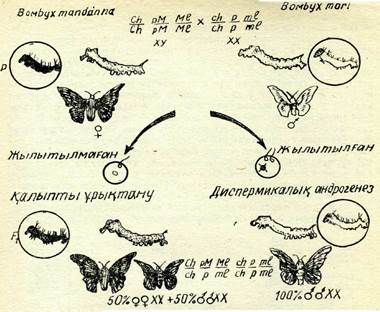
53-сурет. Тұқым қуалауды ядро мен цитоплазманың салыстырмалы қызметін анықтауға мүмкіндік беретін тәжірибе схемасы (жібек көбелегіне жылумен әсер ету әдісі арқылы диплодты андроген особьтар алу): сһ —личинканың ңара түсі; сһ —личинканың саргыш ңоңыр түсі; рМ —көбелектің нүцгірт түсі; р— аң түсі; МІ — жүлдыз ңүртының сүр түсі; ті — ақ түсі.
2. ХРОМОСОМАСЫЗ, ЯҒНИ ЦИТОПЛАЗМАЛЫҚ ТҰҚЫМ ҚУАЛАУ
Пластидтік тұқым қуалау. Пластидтер өсімдік организмінде углеводтарды синтездеуші ерекше лаборатория болып табылады (1-тарауды қараңыз). Оларда ДНҚ бар, бөліну арқылы көбейе-ді және жас клеткаларға таралады. Клетка пластидтерінің жи-ынтығы тұқым қуалаушы информацияны бере алатын құрылым ретінде пластидом деп аталған болатын.
Пластидтік тұқым қуалау жөнінде алғашқы мәліметтерді ге-нетиканың дамуының бастапқы кезінде (1909 ж.) Э. Баур мен К. Корренс хабарлады. Мысалы, Қорренс түн сұлуы (МігаЬіііа ]а1ара) атты өсімдіктегі ала жапырақтылықтың тұқым қуалау-ын зерттеді. Осы түрде ала жапырақты өсімдіктер кездеседі, олардың өсу нүктелерінде әр түрлі клеткалар топтары: қалыпты пластидтілері және хлорофилл түзуге қабілетсіз пластидтілері бар. Осының салдарынан өсімдіктерде кейде таза жасыл түсті немесе аппақ түсті бұтақтар түзіледі. Ақ түсті бұтақтан алынған дәыдер тіршілік етуге қабілетсіз өскіндер береді, өйткені ондай өскінде фотосинтез процесі жүрмейді.
Ала жапырақты бүтақтардағы гүлдерді жасыл түсті жапырақты бұтақтардан алынған гүл тозаңдарымен тозаңдандырғанда және керісінше шағылыстырғанда әр түрлі нәтиже алынады. Бірінші жағдайда ('? ала жапырақты X сГ жасыл жапы-рақты) будан өсімдіктер ала жапырақты, жасыл жапырақты не-месе ақ жапырақты (өледі) болады. Керісінше шағылыстырған кезде ($ жасыл жапырақтыХ <? ала жапырақты) үрпақтағы барлық өсімдіктер жасыл жапырақты болады. Жасыл бұтақтардың гүлдері тек қана жасыл түсті үрпақ береді, ал ақ түсті бұтақтардың гүлдері қандай өсімдік тозаңымен тозаңдандырғанына қарамай ақ түсті үрпақ береді (тіршілік ете алмайды) (54-сурет). Осының негізінде мынадай қорытынды жасалды: ала жапырақтылықтың тұқым қуалауы клетка бөлінуі кезінде екі түрлі пластидтің — жасыл түсті және бояусыз пластидтердің — берілуі және таралуымен байланысты, бүл кезде пластидтер жұмыртқа клеткасымен беріледі, осының нәтижесінде тұқым қуалау аналық линия бойынша іске
асады. Алайда кейбір жағ-
54-сурет.өсімдігінде ала жапырақтылықтың тұқым қуалауы. Аналық форма ретінде мынадай жапырақтары бар өсімдіктер алынған:3— жасыл түсті; 2 — ала. 
55-сурет Клетка бөлінуі кезінде ақ және жасыл түсті пластидтердің кездей-соқ бөліну схемасы:
1— екі түрлі клетканың түзілуі, оның біреуі (а) аң түсті, ал екіншісі (б) ала түсті учяпке береді. 2 —екі түрлі клеткяның түзілуі, оның біреуі (в) жасыл түсті, ал екіншісі (г) ала түсті учаске береді; АБ және ВГ —клеткалардың бөліну сызыңтары. дайларда, мысалы қазтамақ өсімдігінде пластидтер жұмыртқа клеткасы арқылы ғана беріліп қоймайды, цитоплазмасы бар спермия арқылы да беріледі. Бұл кезде ала жапырақтылык ана-лық қана емес, аталық линиямен де тұқым қуалайды, яғни екі ата-аналық жолмен тұқым қуалайды.
Екі түрлі пластиді бар зиготадан өсімдіктердің ақ түсті не-месе жасыл түсті бөліктерінің дамуы әр түрлі пластидтердің өзін-өзі өндіру жылдамдығы және клеткалық бөлінулер бары-сында олардың таралуымен анықталады. Тек жасыл түсті плас-тидтер алған клеткалар тканьдердің жасыл түсті учаскелерін береді, ал боялмаған пластидтері ғана бар клеткалардан ақ түс-ті учаскелер пайда болады. Егер екі сортты пластиді бар клетка АВ сызығы бойынша бөлінсе (55-сурет), екі түрлі учаске бере-тін (ақ түсті— а және ала түсті— б) екі клетка түзіледі; ВГ сы-зығы бойынша бөлінгенде жасыл түсті және ала түсті беліктер (е, г) пайда болады.
Р. Сэджер 1972 жылы бір клеткалы балдыр хламидомонада (Сіатісіотопаз геіпһагсіі) хлоропластардың генетикалық аппа-раты сақиналанған тіркесу тобына үйымдасқандығын анықта-ды. Бактерияларда да генетикалық аппарат осындай қүрылысты болады. Митохондриялар арқылы тұқым қуалау. Митохондриялар-дың тыныс алу процесіне тікелей қатысы бар (1-тарауды қара-ңыз). Олар беліну арқылы өзін-өзі өндіруге қабілетті. Олардан ДНК табылды. Бәліну нәтижесінде жас клеткалар аналық клетка митохондрияларының жартысын алады
 |
| Гиллоидты спорал&р Дип/гоид |
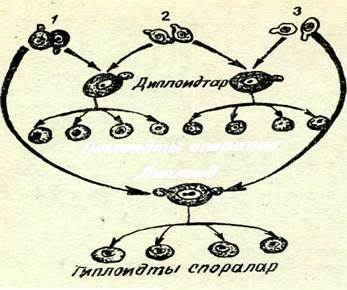
56-сурет. Ашытқы саңырауқұлақтардыңжырап айрылушы және вегетативті ергежейлі штаммаларының генетикалық анализінің схемасы.
і— ңалынты; 2-ажырап айрылу кездегісі; 3 — вегетатив-тісі.
Кейбір саңырауқұлақтарда (ашытқы саңырауқұлақтарда, нейроспорада) тыныс алу процесінің жеткіліксіз екендігі табылды, ол жағдай митохондриялар қызметіндегі тұқым қуалайтын қайтымсыз өзгеріс-цитохромоксидаза активтігі жоғалғандығы-мен қамтамасыз етілген.
Б. Эфрусси Зассһаготусез сегеүізіае ашытқы саңырауқұлақтың спонтанды пайда болатын тыныс алу процесі жеткіліксіз ергежейлі колониялар түзетін штаммаларын тапты. Колониялар гаплоидты саңырауқұлақтардың вегетативті көбеюі кезінде түзі-летіндіктен, олар вегетативті ергежейлілер деп аталды. Вегета-тивті ергежейлі колониялармен қатар, фенотипі — өсуі және тыныстану жетімсіздігі бойынша біріншіге ұқсас форма табыл-ды, бірақ ол ядролық бір генмен анықталған секілді ергежейлік белгісі бойынша ажырау берді. Бұл форма ажыраушы ергежей-лі штамма деп аталды.
Вегетативті және ажыраушы ергежейлі штаммаларға (56-су-рет) генетикалық анализ жасау мынаны көрсетті: ажыраушы ергежейліліктіқ фенотипі ядролық генмен анықталады, өйткені шағылыстыру кезінде аскоспоралардың ажырауы 1: 1 қатынасында болады. Вегетативті ергежейлілер мен қалыпты ашытқы саңырауқұ.лақтарды шағылыстырғанда алынған диплоидты зи-гота (мұнда қалыпты форманың митохондриялары бар) ажырау бермейді, аскоспоралардан ұсақ колониялар пайдаболмайды. Демек, бұл формалардың геномдары бірдей, тек цитоплазмасында ғана айырма бар. Мейозда цитоплазма типімен ажырау болмайды. Бұл экспериментте цитоплазмалық тұқым қуалау фактісі айқын байқалады.
Ашытқы саңырауқұлақтардан тыныс алу жеткіліксіздігінің ьтұқым қуалау арқылы берілуінде митохондриялардың ролін көрсететін тікелей дәлел алынды. Клетка қабықтары жоқ вегетативті ергежейлілер қалыпты саңырауқұлақтардың оқшауланған митохондриялары бар жағдайда өсірілді. Осының нәтижесінде түзілген колониялардың бір бөлігінің (2—2,5%) үлкендігі қалыптағыдай болды. Бүл фактіні мынадай болжау жасап түсіндіуге болады: «қалыпты» митохондриялар вегетативті ергежейлілердің клеткаларына түсіп, олардың тыныс системасындағы кемістігін жойды, бөліну барысында клеткадан клеткаға беріле отырып, қалыпты колониялар түзілуге себеп болды.
I Ңитоплазмалық аталық стерильділігі. Цитоплазмалық түқым қуалаудың ең айқын да нақты мысалының біреуі — цитоплазмалық аталық стерильділік (ЦМС) қүбылысы ол жүгері, жуақызылша, зығыр сияқты көптеген өсімдіктерден табылды. Жүгерідегі цитоплазмалық аталық стерильділігін 30-жылдары СССР-де М. И. Хаджинов және АҚШ-та М. Родс бір мезгілде ашты. Жүгері бір үйлі өсімдік, оның аналық гүлдері собыққа,аталық гүлдері шашаққа жиналған. Жүгерінің кейбір сорттарынан шашақтарында көбінесе ішінде тозаңы жоқ, бос, кейде жетілмеген стерильді тозаңдары болатын жетілмеген тозаңдықтары бар өсімдіктер табылды. Бүл белгі цитоплазманың ерекшеліктерімен анықталатын болып шықты. Аталық стерильділігі бар өсімдіктерді басқа өсімдіктердің қалыпты тозаңымен тозаңдандырса,көпшілік жағдайда үрпағы стерильді тозаңы бар өсімдіктер береді. Бүл шағылыстыруды бірнеше үрпақ бойы қайталағанда, аталық стерильділік белгі аналық линия арқылы беріле отырып, жоғалып кетпейді. Тіпті стерильді тозақы бар өсімдіктердің 10 жұп хромосомаларын фертильді тозаңы бар өсімдіктердің хромосомаларымен ауыстырғанда да, аталық стерильділік сақталады. Осы белгінің цитоплазма арқылы түқым қуалауына бүл сенімді дәлел болды. Тозаңның стерильділігін қамтамасыз ететін цитоплазма(стерильді цитоплазма) белгісімен, фертильді тозаңы бар өсімдіктердің цитоплазмасы (қалыпты цитоплазма) белгісімен көрсетіледі.
Стерильді цитоплазманың қасиетіне өсімдіктің генотипі бел-гілі дәрежеде әсер ете алатыны анықталды. Өсімдік генотипінде рецессивті ген гомозиготалы күйінде болса ғана цитоплаз-ма тозаңның стерильді болуын қамтамасыз ете алады.. Егер осы ген доминантты аллель Я күйінде болса, онда өсімдігінің немесе өсімдігінің тозаңы калыпта-ғыдай болады. Сонымен аллелі тозаңның фертильділігін қал-пына келтіруші болып табылады. Демек, өсімдіктердің бәрінде фертильді бола алады,өсімдігінің ғана тозаңдары толық стерильді болады.
Шағылыстыруларын көп қайтала-ғанда да әр уақытта ұрпақтарындағы тозаң толық стерильді болады. Тек шағылыс-тырулары кезінде ғана цитоплазмасының болғанына қарамай, барлық өсімдіктерінде қалыпты
тозаңы бар үрпақ алы- 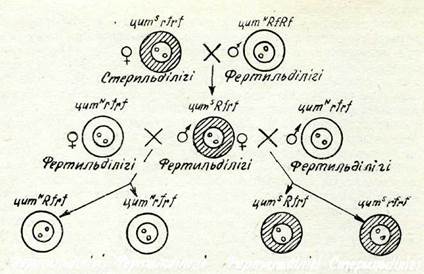
Фертильділігі Стермльаілкі 57-сурет. Цитоплазмалық аталық стерильділігінің тұқым қуалау схемасы: цитоплазма стерильді; К/—ген — тозаңның фертиль-ділігін ңалпына келтіруші.нуы мүмкін (57-сурет). цитоплазманың құрылымы мен ерекшелігін өзгертпейтінін, тек оның әсерінің көрінуін те-жейтіндігін тағы да ерекше атап өтуді қажет етеді.
3. ЦИТОПЛАЗМАНЫҢ ПРЕДЕТЕРМИНАЦИЯСЫ
Цитоплазма предетерминациясының мағынасы. Бірқатар жағдайда белгілердің тұқым қуалауы организмнің жеке басы-ның даму процесінде, немесе сыртқы орта факторлары әсерінен (онтогенездік немесе фенотиптік предетерминация) немесе гено-тип әсерінен (генотиптік предетерминация). пайда болатын ци-топлазманың ерекшеліктерімен байланысты болады.
Онтогенездік предетерминация. Бұл жағдайда кейбір белгі-лердің аналық линия бойынша тұқым қуалауын белгілі бір сыртқы факторлардың әсерінен пайда болатын цитоплазмадағы өзгерістер қамтамасыз етеді. Әдетте мұндай өзгерістер тұрақты емес және бірнеше ұрпақтан кейін біртіндеп жойылып кетеді де, бастапқы типке қайтып оралады. Мысалы, шаншардың құрғашысының жұмыртқасына ұрықтанғанға дейін жоғары температурамен әсер ету, олардың үрпағының денелеріндегі реңді өзгертуге әкеледі. Келесі ұрпақтар қалыпты температура жағдайында көбейгенде бүл өзгеріс біртіндеп жойылады. Ал насекомның еркегіне жоғары температурамен әсер етіп, үрғашыларын қалыпты жағдайда өсірсе, мүндай қүбылыс байқалмайды.
Организмдерді бастапқы жағдайларға көшіргенде оның келесі ұрпақтарында біртіндеп жоғала бастайтын мүндай өзгерістерді созылыщы модификация деп атайды. Олардың механизмі осы уақытқа дейін анықталған жоқ. Өзгерістерді туғызған фак-торлар сақталса, созылыңқы модификациялар бірнеше ұрпақтар бойы үнемі көріне береді; ал ондай әсер ететін факторлар ■болмаса, біртіндеп бастапқы қалыпқа түседі (23-тарауды қара-ңыз).
Генотиптік предетерминация. Аналық организм генотипінщ әсерінен цитоплазма қасиеттерінін, предетерминациялану фактілері көңіл аударарлық құбылыс. Тұщы суда тіршілік ететін Ілтпеа ұлуы қабыршағының бұралу бағыты бұған айқын мы-сал бола алады: олардың кейбіреулері гермафродиттер болып табылады және өзін-өзі ұрықтандырып та, өзара шағылысып та көбейе алады. Қабыршақтарының бұралуы екі түрлі болады: сағат стрелкасына қарсы — сол жаққа қарай бұралғандар және сағат стрелкасы бойынша оң жаққа бұралғандар. Қабыршақ-тың бұралу бағытын бір жүп аллель анықтайды: оң жаққа қарай бүралу О, сол жаққа қарай бүралуға сі доминанттылық көрсетеді.
Реципрокты шағылыстырулар кезінде генотиптері бірдей бу-дандар фенотипі бойынша айрылады. шағылыстыруында барлық буданды особьтар аналық типтес—қабыршақ-тары оң жаққа қарай бүрылған боладышағылыс-тыруында да үрпақ қабыршағының бүралуы аналық типтес, яғни сол жаққа қарай бүралған болады (58-сурет).
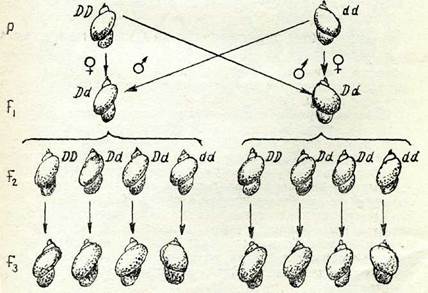
58-сурет. Іітпеа моллюска қабыршағы бұралуының тұқым қуалануында цитоплазма ролі: В —оң жавда ңарай бұралуын баоңяратын ген, й —сол жаңда ңарай бұралуын басңаратын ген.
Фенотиптері бойынша Ғ\ будандарының (аналық формалар) айырмашылықтары болғанмен шағылыстырудың екеуінен де алынған гетерозиготалы формалар Ғ\ (Ш) өзін-езі ұрықтандырған кездедегі барлық ұрпақтардың қабыршағы оң жақ-қа қарай бұрылған болады,әр особыңың ұрпақтарын же-келеп зерттеген кезде 1/4 семьялар сол жаққа қарай бұралған, ал 3/4 бөлігі оң жаққа қарай бұралған болып шықты. Осы жұп белгілер бойынша мендельдік ең карапайым ажырау 3: 1 Ғ2 кезінде емес, тек Ғз-те ғана байқалды. Мұндай типті тұқым қуалау кезінде ұрпақтар фенотипі өздері дамитын зиготалар гено типіне емес шешесінің генотипіне сәйкес келеді, осы белгі жұмыртқа цитоплазмасының дамуы процесінде аналық организмнің генотипімен алдын ала анықталады деп жорамал жасағанда ғана көрінетін болады. Қаралған тұқым қуалау типі шын мағы-насында аналық жолмен тұқым қуалау болып табылады.
Анықталғандай қабыршақ бұралуының бағыты ұрықтанған жұмыртқаның спиральдана бөлшектену сипатымен анықталады, яғни бластомерлердің спираль бойымен оң жақта немесе сол жақта орналасуымен бұл өз кезегінше екінші рет бөлшектене бөліну кезіндегі ұршықтың бағытталуына тәуелді.
Қарастырылған бұл мысал, дәлірек айтқанда, цитоплазмалық тұқым қуалауға тікелей қатысты емес, өйткені мұнда цитоплазманың элементтерімен емес, хромосомалық гендердің әсері-мен анықталған, яғни жұмыртқа клеткасының цитоплазмасын тіпті ұрықтануға дейін өзгертетін хромосомалық тұқым қуалау механизмі әсер етеді.
4. ИНФЕКЦИЯ ЖӘНЕ ЭНДОСИМБИОНТТАР АРҚЫЛЫ ТҰҚЫМ ҚУАЛАУ
Өзін-өзі өндіре алатын қасиеттері бар әр түрлі симбионттар цитоплазма арқылы берілуі мүмкін және осының нәтижесінде цитоплазмалық тұқым қуалағандай алдамшы кәрініс (имита-ция) бере алады.
Инфекция арқылы тұқым қуалау. Тышқандардың сүт безде-рінде рак ауруы дамуына бейім тұқым қуалайтын линия бо-лады, ауруға ондай бейімділік тек үрпағын емізу кезінде ғана аналық линия арқылы тұқым қуалайды. Егер ракты линиялар-дан аналарына — сүтпен асыраушыларға ракты емес линиялар-дан тышқан балаларын салып қойса, онда тышқанның мұндай балалары да сүт бездерінің рак ауруына бейім болып еседі. Егер ракты линиядан туған тышқан балаларын туғаннан бас-тап қалыпты тышқандар емізсе, онда тышқан балалары ауырмайтын болады. Сонымен катерлі ісікті бүл жағдайда ана сүтіарқылы берілетін инфекция туғызады. Бүл инфекциялық агент сүт факторы деп аталды. Онда вирустық табиғат бар екені анықталды.
Эндосимбионттар арқылы тұқым қуалау. Рагатесіит аигеііа инфузорияларының кейбір линияларының цитоплазмасында каппа-бөлшектер деп аталатын арнаулы бәлшектер болады және оны ол өзі жүрген ортаға беліп шығара алады. Каппа бөл-
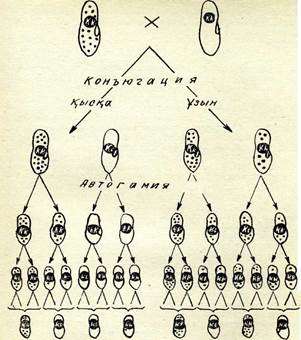
59-сурет. Инфузорияда К гені аллельдерінің және каппа-бөлшектерінщ тұқым қуалау схемасы. Қаппа-бөлшектері қара нүктемен белгіленген.
шектердің өз иелеріне («өлтіргіштер») ол әсер етпейді, ал бас-қа линиялардың парамецріялары (сезімталдар) ондай заттардың әсерінен өледі.
Каппа бөлшектері парамециялар үшін эндосимбионттар бо-лып табылатын бактерия (СаесіоЬасіег іаепіозрігоііз) екені жақында анықталды. Ондай бактериялардың өздеріне тән белок лентасы бар, ол ленталарда бактерия симбионттары-фагтар орналасады. Сонымен мұнда өзіне тән үш жақты симбиоз бар; инфузория—бактерия—фаг.
Каппа-бөлшектер сезімтал инфузорияның ас қорытушы ва-куоліне түскеннен кейін бактерияның белок лентасы жазылады.Фагтардың тіршілік әрекеті нәтижесінде белок табиғатты бір заттар түзіледі, сірә, инфузория өліміне осы зат себепкер болса керек.
Каппа-бөлшектердің цитоплазмада сақталуын және олардың «өлтіргіш» инфузориялардың бөліп шығаруын доминантты ген /(бақылайды; оның рецессивті аллелі к олардың сақталуына әсер етпейді. «Өлтіргіш» инфузориялар тікелей бөлінгенде үнемі осындай «өлтіргіш» қасиеттері бар клон береді. Тиісті жағдай-ларда шағылыстыруға мүмкіндік туады, яғни «өлтіргіш» және сезімтал инфузорияның екі клеткасы конъюгацияланады.
59-суретте К және к аллельдерінің түқым қуалау және кап-па-белшектердің таралу схемасы берілген. Бастапқы формалар гомозиготалар (КК және кк) болып табылады, &л эксконъюгант-тар — гетерозиготалылар (Кк). Эксконъюганттардың келесі автогамиялары кезінде микронуклеус пісіп-жетілуінің екі бөлінуі жүреді. Пайда болған гаплоидты терт ядроның үшеуі дегенера-цияға үшырайды, ал қалған бір ядро митоздық жолмен бөлі-неді. Осының нәтижесінде гаплоидты екі пронуклеус түзіледі де, сонан соң қүйылысып қосылады, сөйтіп дпплоидты және міндет-ті түрде гомозиготалы микронуклеус (қарастырылып отырған мысалда КК немесе кк) пайда болады. Эксконъюганттар авто гамиясы нәтижесінде генотипі бойынша 1: 1 қатынасында ажырау байқалады, «өлтіргіш» (КК) мен сезімтал (кк) каппа-белшектерінің таралуы ата-аналық клеткалар конъюгациясы-ның үзақтығына тәуелді болады. Егер конъюгация қысқа мер-зімді ғана болса және тек микронуклеустер ғана алмасса, ал эксконъюганттар цитоплазмасын алмастыруға үлгіре алмаса, онда каппа-белшектер сезімтал әріптесінің цитоплазмасына ен-бейді және бастапқы клетканың тек езінде ғана қалады. Егер конъюгация жеткілікті үзақ уақытқа созылса, онда сезімтал клеткадан шығатын эксконъюгант К генін ғана қабылдап алып қоймайды, өзі генотипі бойынша Кк болады, цитоплазмасында да каппа-бөлшектер болады. Келесі бәлінулерде бұл парамеция «өлтіргіш» клон береді. Каппа-бәлшектер инфузория цитоплаз-масында көбейеді және К аллелі болса ғана сақталады. Егер олар сезімтал клетканың (кк) цитоплазмасына түсетін болса, онда олар көбеймейді және ары қарай бәлінгенде біртіндеп «сүйылады», ақырында жойылады.
Біз қарастырған мысалдар мынаны көрсетеді: кейде цито-илазмада эндосимбионттар бола алатын, өзін-өзі өндіруге қабі-летті, бірқатар қасиеттерді аналық линия арқылы бере алатын бактериялар кездеседі. Бірақ бүл симбионттар шын мағынасын-да тірі клетканың бөлінбес элементтері болмайды.
5. ХРОМОСОМАСЫЗ ТҰҚЫМ ҚУАЛАУДЫҢ ГЕНЕТИКАЛЫҚ АНАЛИЗІ
Осы уақытқа дейін біз генотип деген терминді хромосома-ларға жинақталып орналасқан гендер системасы деп қана біліп келдік. Ал цитоплазмада және оның органоидтарында жинақталған тұқым қуалаушы факторлар плазмотип немесе плазмон деген терминмен белгіленеді. Генотип деген ұғымды кеңейтіп, ол ұғымға хромосомалық гендер системасын (геном) да, цитоплаз-малық тұқым қуалау системасын (плазмон) да енгізу дұрыс бо-лар еді. Хромосомалық тұқым қуалаудың бірлігіне (генге) сәйкес келетін
 |
цитоплазмалық тұқым қуалаудың кез келген бірлігін плазмоген деп атайды.
Клетканың генетикалық аппаратының системасы Геном — Хромосомалар — Гендер
генетикалық материалы
Генотиптің кеңейген мағынасы өзіне клетканың бүкіл гене-тикалық системасын енгізуі керек, сонда ғана тұқым қуалау жөніндегі осы заманғы ұғымға өте сәйкес болады.
Хромосомасыз тұқым қуалаудың генетикалық анализі. Ци-топлазмалық тұқым қуалауға генетикалық анализ жасау үшін анығында, клетка органоидтарының биохимиясы және морфоло-гиясы, олардың клетка цикліндегі санының тұрақтылығы неме-се өзгеру заңдылықтары, олардың өзін-өзі ендірулері мен қыз-меті жөнінде цитологиялық және биохимиялық дәл мәліметтер қажет. Цитоплазмалық тұқым қуалауды генетикалық әдістер арқылы табысты түрде зерттеу үшін, ең алдымен, цитоплазманың предетерминациясын және инфекцияның барлық типтерін шығарып тастау қажет. Цитоплазманың әр түрлі органоидтарымен байланысқан және айқын көрінетін белгілері бар формалар алу қажет.
Хромосомасыз тұқым қуалауға генётикалық анализ жасаудың нақты мысалын кдрастырайық. А. Срб нейроспора түрлерінің біреуінен (Ы. Зііорһуііа) бірнеше форма алып, оларды 5^ деп атады. Олар споралары мен конидияларының баяу өсуімен сипатталады, бірақ кейінірек олардың есуі қалыпты жылдамдықпен өткен. Цитохромдарына анализ жасау олардың жабайы типтен айырмашылығы жоқ екенін көрсетті, демек баяулап өсу тыныс алудың бүзылуымен байланысты емес.
5,§"-формаларды жабайы типпен (Ы) реципрокты шағылыс-тыру аналық түқым қуалау болатынын көрсетті. Беккростардың бірнеше үрпақтарын (жиырмаға дейін) зерт-теу аналық хромосомаларды аталық хромосомалармен толық алмастырған кездің өзінде 5§" белгісі өзгеріссіз аналық жолмен түқым қуалайтынын көрсетті.
 |
5^-белгінің хромосома гендерімен тіркесуін сынау оның ней-роспорадағы жеті хромосома гендерінің ешқайсысымен де тір-
кеспегенін көрсетті. Тетрадалық анализдің көмегімен бойынша ажыраудың жоқтығы хромосома гендері бойынша ажырауға әсер етпейтінін анықтады. Сонымен, 5-ның цитоплазмалық тұқым қуалау сипатын ядродағы кандай да болмасын бұзылумен мейоз-дың ерекшеліктерімен түсіндіруге болмайды.
Осыдан кейін цитоплазма-лар араластырылды — нейро-спораның екі формасының гитерін эксперименттік жолмен қосты: оның біреуінде қалып-ты цитоплазмасы және хромосомалық маркер а (ақ мице-лий), екіншісінде өсудің баяу-лауын анықтайтын 5д цито-плазмасы және боялған мицелий беретін аллелі а+ бар. Кешіпей, гифтердің құйылысып қосылғанын микроскоп астын-60-сурет. Әр түрлі мицелийлердің да көргеннен кейін, осы учаскеде пайда болған ядроларьг ядролық факторлар бойынша кй- бар өскіндердің бәрі кесіп алы роспораның ажырап айрылуы:нып, жекелеген пробиркаларға
1— мицелийлердің өзара ңұйылысуы;салынды. өсіп шыққан мицо-құрамында аралас цитоплазмасы бар,гетерокарион мицелийінің учаскесі; 3—ЛИИЛбрДеН КОНИДИЯЛар (оларды бөліп өсірудің нәтижесі. Цитосының көбею ілазмальқ факторлар:;өсетіндері және 8—баяу есетіндері.жиналды және анализ - ~™т;::іт2::т%&. *«■
машн мицелий. арасында Л/а мен г>§а+-къщ бастапқы үйлесімдері ғана емес, сонымен қатар 5£ а мен N а+-ның жаңа үйлесімдері табылды (60-сурет). ІДитоплазмалық тұқым қуалаудың дискреттілігін дәлелдейтін цитоплазмалық фактор бойыныша ажырау табылды және мұндай ажырау хромосома гендерінің таралуынан тәуелсіз болып шықты.Сонымен, баяндалған фактілердің негізінде үрпақтар ара-сындағы материалдық және қызметтік сабақтастылық клетканың өзін-өзі өндіре алатын — ядролық және цитоплазмалық құрылымдарының бәрімен қамтамасыз етіледі деп есептеуге бола-ды. Хромосомалық тұқым қуалау сияқты цитоплазмалық тұқым қуалау да дискретті. Ядролық тұқым қуалаудан оның айырмасы — оаналық жолмен тұқым қуалаиды және онда ажырау-дың тұрақты сандық зандылықтары болмайды.
Ядролық гендер сияқты, цитоплазмалық гендер де тұрақтыдығьмен гетерозиготалы жағдайда өзгермеушілігімен, аллельдерінің ажырауымен сипатталады, оларға генотип және фенотип жөніндегі ұғымдар қолданылады.
11-тарау. ТҰҚЫМ ҚУАЛАУДЫҢ НЕГІЗГІ ЗАҢДАРЫ ЖӘНЕ ТҰҚЫМ ҚУАЛАУШЫЛЫҚ ПРИНЦИПТЕРІ
Тұқым қуалаушылық және тұқым қуалау кейде ажырата алмайтын екі түрлі құбылыс. Тұқым щуалау — көбею процесінде организмнің тұқым қуалау арқылы анықталатын қасиеттері мен белгілерінің, бастамаларының, ата-анадан ұрпаққа берілу про-десі. Тұқым қуалаушыльіКі деп клетка құрылымдарымен тұтас организмнің ұрпақтар арасындағы материалдық және қызметтік сабақтастылықты қамтамасыз ету қасиетін айтады. Тұқым қуа-лау және тұқым қуалаушылықтың негізінде тұқым қуалай өзін-өзі өндіре алатын маңызды дәл құрылымдар мен клеткалар бөлінуі кезінде олардың заңды таралуы жатыр.
1 МЕНДЕЛЬДЩ ТҰҚЫМ ҚУАЛАУ ЗАҢДАРЫ ЖӘНЕ ОЛАРДАН ШЫҒАТЫН ТҰҚЫМ ҚУАЛАУШЫЛЫҚТЫҢ ПРИНЦИПТЕРІ
Бірінші ұрпақ будандарының бірдей болуы, ажырау және белгілердің тәуелсіз комбинациялануы жөніндегі Мендель аш-қан заңдылықтар, сол сияқты Морган ашқан тіркесу және жыныспен тіркескен белгілердіц тұқым қуалауы тұқым қуалаушылыққа жатпайды, тұқым қуалау заңдылықтарына жатады.
Оларды ашудағы Мендельдің табысы онын, жекелеген жұп белгілерге генетикалық анализ әдісін қолдануы қамтамасыз ет-ті. Мынадай құбылыстарды ашу арқылы Мендель шын мағына-сында генетиканың ғылыми негізін қалады:
1. Әр белгі жыныс клеткалары арқылы берілетін тұқым қуалайтын фактормен анықталады; қазіргі заманғы түсінік бойынша бұл бастамалар гендерге сәйкес келеді.
2. Үрпақтар қатарында гендер өздеріне тән ерекшеліктерін жоғалтпай және езгермей таза күйінде сақталады, яғни ген салыстьірмалы түрде түрақты.
3. Түқым қуалайтын бастамалар жүп болады: біреуі—анасы нікі, екіншісі әкесінікі; олардың біреуі доминантты, екіншісі рецессивті болуы мүмкін; бүл ереже аллелизм принципін ашуға сәйкес келеді, ол принцип бойынша генде әрқашан ең кем дегенде екі аллель болады.
4. Екі жыныс та өздерінін, түқым қуалайтын қасиеттерін ұрпақтарына беруде бірдей мөлшерде қатысады.
5. Гендердің саны жыныс клеткаларында екі есе азаяды; бұл ереже мейоздың болатындығын генетикалық алдын ала болжау болып табылады.
Айтылғандардың негізінде Мендельдің тікелей өзі тұжырымдаған және тұқым қуалау процесіне қатысы бар заңдарды және Мендель жұмысынан келіп шығатын тұқым қуалаушылық прин-циптерінен ажырата білудің пайдалы екені түсінікті.
Тұкым қуалау заңдарына: бірінші ұрпақ будандарының бір-дейлілік заңы, будан ұрпағында тұқым қуалайтын белгілердің ажырауы және тұқым қуалайтын белгілердің тәуелсіз комбина-циялану заңы жатады. Бұл заңдар жынысты көбеюде тұқым қуалайтын информацияның ұрпақтарға берілу процесін қамтып көрсетеді.
Тұқым қуалаушылық принциптерінде басқа да мағына бола-ды, оларды төмендегідей тұжырымдауға болады:
1. Белгілердің дискретті (генді) тұқым қуалай детермина циялануы.
2. Тұқым қуалайтын бірліктің—геннің салыстырмалы тұрақтылығы.
3. Геннің аллельдік күйі (доминанттылық және рецессивтілік).
Мендельдің тұқым қуалау заңы және олардан келіп шығатын тұқым қуалаушылық принциптері генетиканың негізгі мазмұны болып табылады. Олардың ашылуы осы заманғы табиғаттану ғылымына тіршілік процестерінің &#








