2015-09-06
2015-09-06 344
344Крім соматичної гібридизації і клітинної селекції є ще один метод створення in vitro покращених форм господарсько цінних рослин – це генетична трансформація.
Термін трансформація походить від латинського transformatio – перетворення і означає зміни в геномі та спадкових властивостей організму.
Генетична трансформація досягається шляхом проникнення крізь канали клітинної мембрани екзогенної ДНК і взаємодії останньої з клітинним геномом.
На відміну від соматичної гібридизації і клітинної селекції генетичну трансформацію розглядають як спосіб отримання більш обмежених і специфічних генетичних змін. У багатьох випадках окрема важлива у сільськогосподарському відношенні ознака, що контролюється одним або невеликою кількістю генів, може бути змінена шляхом трансформації, прицьому більша частина геному залишається без змін. Технологія рекомбінантних ДНК дозволяє виділити у чистому вигляді і в достатній кількості будь-який ген і при наявності вектора або іншим способом вбудувати цей ген до складу хромосомної ДНК і добитися його експресії. Використання цієї технології робить пошук цілеспрямованим і значно розширює можливості маніпулювання генетичним апаратом при скороченні часу отримання нових форм рослин.
В усьому світі проводяться дослідження по визначенню, виділенню, клонуванню і введенню в рослини генів, які відповідають за стійкість до холоду, посухи, засолення, тобто до стресових впливів оточуючого середовища, за стійкість до шкідників, гербіцидів і пестицидів, хвороб тощо. Той факт, що геном рослин вивчений гірше, ніж геном ссавців значно ускладнює такі дослідження. На сьогодні розроблено багато підходів до проблеми трансформації, однак з практичної точки зору відносно простий і відтворюваний метод базується на використанні Agrobacterium tumеfaciens.
A.tumеfaciens – це ґрунтова бактерія із родини Rhizobiaceae, яка здатна, поселяючись на раневих поверхнях рослин викликати утворення пухлин. Власне пухлинотворним агентом є плазміда, частина якої – Т-ДНК (від англ. transferred DNA), переноситься в хромосоми клітин рослини (Рис. 7). Плазміда – невелика кільцева молекула ДНК, здатна до стабільного, не зв’язаного з хромосомами існування і автономної реплікації (біосинтез дочірніх ДНК). Плазміди можуть вбудовуватися у хромосоми. Плазміди локалізовані в цитоплазмі бактеріальних клітин і клітинах деяких дріжджів. Після трансформації рослинні клітини починають швидко і неорганізовано рости і синтезувати особливі речовини – опіни (похідні амінокислот і цукрів), які бактерії використовують як джерело азоту і вуглецю. Тобто утворення пухлин супроводжується перебудовою метаболізму клітин, і вони починають синтезувати сполуки, необхідні лише для бактерій.
Раніше припускали, що A. tumеfaciens вражає саме пошкодженні рослини внаслідок руйнування клітинної стінки і усунення фізичного бар’єру, який ускладнює проникнення бактерій у клітину. Однак сьогодні вважається, що індуктором генетичної трансформації у агробактерій є фенольні сполуки, ацетосирингон і гідроксіацетосирингон, які виділяють рослини при пораненнях.Плазміди, які викликають пухлини називаються Ті-плазмідами (від англійського tumor inducing – індукуючі пухлину). Це кільцеві молекули ДНК
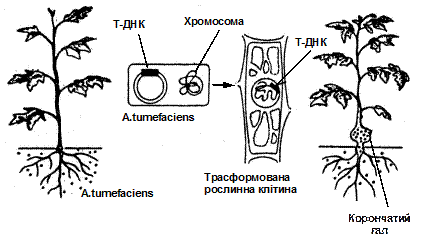
Рис.7. Етапи індукції корончатого галу після інфікування рослини Agrobacterium tumеfaciens.
довжиною близько 200 кб (3-5 % від розміру хромосоми агробактерії), які стабільно зберігаються в агробактеріях при температурі нижче 300С. В бактеріальних клітинах вони реплікуються автономно. Ті-плазміди розрізняють за типом опіну, який синтезується. Найчастіше зустрічаються плазміди, які кодують нопалін або октопін. Вважається, що клітина може містити лише один тип плазміди: або октопінову, або нопалінову.
Генетична карта Ті-плазміди повністю не розшифрована. З’ясовано тільки 4 ділянки: дві онкогенні (Т-область і vir -область) і дві ділянки (OR1, CON), які визначають морфологію пухлин, забезпечують реплікацію плазмід та їхню кон’югацію (Рис. 8).Генетичні дослідження показали, що гени, які відповідають за індукцію пухлини, синтез опінів і пригнічення диференціації, розташовані близько один від одного і входять до складу Т-області плазміди, яка вбудовується у хромосому клітини при інфекції. Т-сегмент має довжину близько 20 кб (10 % Ті-плазміди) і вбудовується в різні, очевидно неспецифічні, області хромосоми. На Т-області картовано сім генів, які відповідають за синтез опіну і за пригнічення диференціації клітин. Слід відмітити, що всі гени, які відповідають за перенесення та інтеграцію Т-ДНК, знаходяться не в Т-ДНК, а в області вірулентності (vir -область).
У тих випадках, коли важко регенерувати цілі рослини із окремих клітин, для трансформації використовують Agrobacterium rhizogenes, яка викликає посилене утворення коренів при зараженні рослин. Рослини багатьох видів можуть регенерувати із культури коренів, які індукує ця бактерія. Плазміди, які викликають коренеутворення, називають Ri-плазмідами (від англійського root inducing – індукуючий корені). Корені швидко ростуть у культурі подібно до тканини пухлин і відсутність бактерій не впливає на їхній ріст. З’ясувалося, що клітини корінців також містять опіни, а їх геноми містять по декілька копій Т-ДНК. Ri-плазміди відрізняються від Ті-плазмід тим, що вони є природними 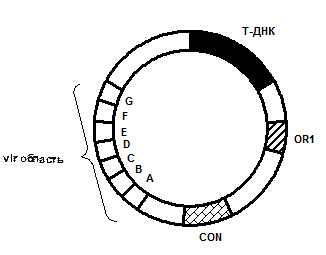
Рис.8. Узагальнена генетична карта Ti-плазміди Agrobacterium tumеfaciens:
vir -область – забезпечує вірулентність;
Т-ДНК – містить ген, який контролює синтез тільки одного із опінів;
OR1 –ділянка реплікації;
CON – ділянка кон’югації.
векторами, після трансформації якими рослинні клітини здатні регенерувати у здорові фертильні рослини. У даний час ці плазміди розглядають як перспективніші вектори для генетичної трансформації рослинних клітин.