 2015-10-16
2015-10-16 19433
19433Динамика популяции — раздел популяционной экологии, изучающий численность особей популяции и механизмы ее регуляции. Жизнь популяции проявляется в ее динамике. К основным признакам динамики относятся:
— численность популяции — общее число особей на данной территории или в данном объеме (например, воды);
— плотность популяции — это среднее число особей на единицу площади или объема;
рождаемость (плодовитость) — число новых особей, появившихся за единицу времени в результате размножения;
— смертность — число особей, погибших за определенный период;
— прирост популяции — разница между рождаемостью и смертностью;
— темп роста — средний прирост за единицу времени. Изучение этих демографических признаков необходимо
для выявления законов жизни популяции, а следовательно, и основ стабильности экосистемы в целом.
Численность популяции никогда не бывает постоянной и зависит от соотношения интенсивности размножения (плодовитости) и смертности. В процессе размножения происходит рост популяции, смертность же приводит к сокращению ее численности.
Рождаемость характеризует частоту появления новых особей в популяции. Различают рождаемость абсолютную и удельную, максимальную и экологическую.
Абсолютная рождаемость — число особей, появившихся в популяции за единицу времени. Удельная рождаемость выражается в числе родившихся особей на число особей в популяции в единицу времени. Например, для популяции человека показателем удельной рождаемости обычно служит число детей, родившихся в год на 1000 человек (таблица 4.2).
Максимальная рождаемость определяется числом самок в популяции и их способностью производить определенное число детенышей в единицу времени (т.е. физиологической плодовитостью). Обычно рождаемость ниже максимальной, т.к. она соответствует сложившимся экологическим условиям и называется экологической.
Таблица 4.2 Рождаемость и смертность в городе Смоленске
| Год | Абсолютная | Удельная | ||
| рождаемость | смертность | рождаемость | смертность | |
| 3550 чел. | 3583 чел. | 10,7 чел. | 10,2 чел. | |
| 2644 чел. | 4480 чел. | 7,7 чел. | 12,7 чел. | |
| 2794 чел. | 4847 чел. | 7,9 чел. | 13,7 чел. |
Экологическая рождаемость дает представление о скорости размножения популяции при фактически сложившихся условиях жизни. Например, взрослая самка трески выметывает миллионы икринок, из которых в среднем доживают до взрослого состояния лишь 2 особи. В результате гибели по разным причинам (болезни, паразиты, хищники и др.) яиц, гусениц и куколок на свет появляется лишь 0,32% взрослых бабочек от числа отложенных яиц.
Численность и плотность популяции зависят также от ее смертности. Смертность популяции — это количество особей, погибших за определенный период. Она, как и плодовитость, изменяется в зависимости от условий среды, возраста и состояния популяции и выражается в процентах к начальной или, чаще, к средней величине ее. У большинства видов смертность в раннем возрасте всегда выше, чем у взрослых особей. У многих рыб до взрослой фазы доживает 1—2% от выметанной икры; у насекомых — 0,3—0,5% от отложенных яиц. Смертность, как и рождаемость, может быть абсолютной и удельной (таблица 4.2).
Выживаемость — средняя для популяции вероятность сохранения особей каждого поколения за определенный промежуток времени. Различают три типа смертности или, как их чаще называют, «кривые выживаемости». Каждый вид имеет свою кривую выживаемости (рис. 11).
Первая кривая — сильно выпуклая. Выпуклость кривой характеризует повышение смертности к концу жизни, остававшуюся до этого низкой. Такой тип кривой характерен для насекомых, личинки которых обитают в почве, воде, древесине или других местах с благоприятными условиями. Он характерен также для проходных рыб, нерестую-щихся один раз в жизни, для многих видов крупных животных и для человека
Вторая кривая — сильно вогнутая. Вогнутость кривой характерна для видов, смертность у которых очень высока на ранних стадиях жизни. Этот тип кривой свойственен большинству растений и животных. Максимальная гибель многих растений происходит в стадии прорастания семян или всходов, а животных — в личиночной фазе или молодом возрасте, например, у устриц, рыб, птиц, многих беспозвоночных.
Третий тип кривой — промежуточный, почти прямая линия, характерна для видов, у которых смертность мало изменяется с возрастом и остается более или менее одина-
ковой в течение всей жизни данной группы. Такая смертность встречается очень редко и только у популяций, постоянно находящихся в оптимальных условиях, например, таких, как гидра пресноводная.
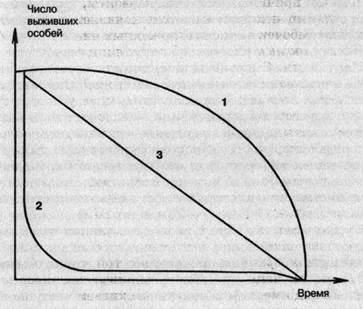
Рис. 11. Три типа кривых выживания: 1 — человек, 2 — рыбы, 3 — гидра
Форма кривой выживания связана со степенью заботы о потомстве и способами защиты молоди. Так, кривые выживания пчел и дроздов, которые заботятся о потомстве, менее вогнуты, чем кривые выживания кузнечиков или сардин, не заботящихся о потомстве.
В замкнутых популяциях (в которых нет миграций) скорость изменения численности определяется только соотношением рождаемости и смертности. Если рождаемость выше смертности, то удельная скорость роста положительная. Если же смертность выше рождаемости, то удельная скорость становится отрицательной и численность популяции начинает убывать. Рождаемость и смертность, т. е. динамика численности, напрямую связаны с возрастной и половой структурами популяции.
Популяция регулирует свою численность и приспосабливается к изменяющимся условиям среды путем обновления и замещения особей. Особи появляются в популяции благодаря рождению и миграции, а исчезают в результате смерти и эмиграции.
При сбалансированной интенсивности рождаемости и смертности формируется стабильная популяция, в которой смертность компенсируется приростом и численность ее, а также ареал поддерживаются на одном уровне.
Популяции, в которых рождаемость превышает смертность и численность популяции растет так быстро, что наступает вспышка массового размножения, называются растущими. Это особенно характерно для мелких животных. Примером может служить увеличение численности популяции колорадского жука (Leptinotarsa decemlineata), быстро расселившегося на территории от Франции до Украины, Белоруссии, Смоленской и Псковской областей. Примером растущей популяции является элодея, завезенная из Америки, появившаяся в 1836 году в Ирландии, и проникшая уже в 1885 году в бассейн Оки. В последние десятилетия наметился рост популяции канареечного вьюрка (Serinus canaria), зеленой пеночки (Phylloscopus trochiloides), чайки обыкновенной (Larus ridibundus), зайца-русака и других видов.
Однако при бурном развитии популяции наступает переуплотнение, что ведет к ухудшению условий существования. А это приводит к резкому возрастанию смертности, в результате чего численность популяции начинает сокращаться. Если смертность превышает рождаемость, популяция становится сокращающейся. Так произошло, например, с элодеей и домовым воробьем (Passer domesticus) в умеренной зоне. В сокращении численности популяций многих животных часто повинен человек; например, таких, как соболь (Martes zibellina), бобр (Castor fiber), зубр (Bison bona-sus), дрофа (Otis tarba) и других. Однако сокращаться безгранично популяция также не может. При определенном уровне численности интенсивность смертности начинает падать, а плодовитость повышается. В итоге сокращающаяся популяция превращается в растущую.
В природе численность популяций всегда испытывает колебания. Амплитуда и период этих колебаний зависят от вида и от условий среды обитания. Различают непериодические (нерегулярные, хаотические) и периодические (регуляр-
ные, циклические) колебания численности популяций. К непериодическим колебаниям численности, а соответственно и плотности популяции, относятся вспышки массового размножения непарного шелкопряда (Ocneria dispar) в южной и юго-восточной частях России в 1879 году, рыжего соснового пилильщика (Neodiprion sertif er) в Ленинградской и Смоленской областях, в Белоруссии с 1958 по 1962 годы. Резкий подъем численности наблюдается у популяций, оказавшихся на новом местообитании. Например, массовое размножение кроликов и разрастание зарослей кактуса опунции в Австралии, колорадского картофельного жука (Leptinotarsa decemlineata) и чайки обыкновенной (Larus ridibundus) в последние годы в Смоленской области.
Периодические колебания повторяются через равные промежутки времени, обычно в течение нескольких лет или одного сезона. Например, циклические изменения с подъемом численности в среднем через 4 года зарегистрированы у леммингов (Dicrostohyx), полярной совы (Nyctea scandiaca) и других животных тундры. Сезонные колебания численности характерны для многих насекомых, мышевидных грызунов, птиц. На периодические колебания численности популяции одним из первых обратил внимание русский генетик С.С. Четвериков (1880—1959), исследовавший изменчивость в природных популяциях. Колебания численности особей, составляющих популяцию, получили название популяционных волн (рис. 12).
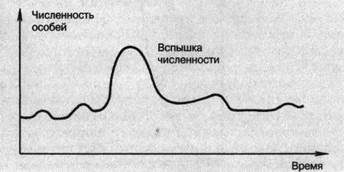
Рис. 12. Популяционные волны
Численность и плотность — основные параметры, которые выражают количественные характеристики популяции как целого.
Каждой популяции свойственен так называемый биотический потенциал — способность к увеличению численности за данный промежуток времени. У разных организмов биотический потенциал не одинаков. Примером организмов с высоким потенциалом размножения являются:
— бактерии Bacillus coli, размножающиеся простым делением каждые 20 минут, которые при наличии условий для реализации биотического потенциала, могли бы освоить все пространство земного шара за 36 часов;
— гриб-дождевик, приносящий до 7,5 млрд спор, уже во втором поколении освоил бы всю Землю.
Крупным организмам с низким потенциалом размножения потребовалось бы для этого несколько десятилетий или столетий.
Таким образом, численность каждого вида при благоприятных условиях способна расти по так называемой экспоненциальной (логарифмической) кривой. Рост численности в геометрической прогрессии называется экспоненциальным ростом. График зависимости численности популяции от времени при экспоненциальном росте представляет собой кривую, напоминающую по форме латинскую букву J, называемую экспонентой. Эта J-образная кривая показывает, что в ходе роста популяции ее численность увеличивается с возрастающей скоростью (рис. 13).
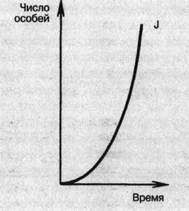
Рис. 13. Экспоненциальная кривая роста популяции
Величина изменения численности за единицу времени называется абсолютной скоростью роста численности. Эта ве-
личина зависит от численности популяции: например, из графика на рис. 14 видно, что чем больше численность, тем больше абсолютная скорость роста. Удельная скорость роста численности — это скорость прироста на единицу особи — эта величина и отражает биотический потенциал.
Экспоненциальный рост в реальных условиях наблюдается, когда популяция растет в условиях избытка ресурсов (пищи, места для размножения), отсутствия конкурентов и не испытывает воздействия неблагоприятных факторов. В лабораторных условиях экспоненциальный рост можно наблюдать у популяций микроорганизмов (дрожжей, бактерий, хлореллы) в начальной фазе их роста. В природе экспоненциальный рост наблюдается при вспышке численности грызунов, саранчи, непарного шелкопряда и других насекомых. Экспоненциально может расти численность популяций, вселенных в новую местность, где у них много пищи и мало врагов. Классическим примером такого роста является рост численности кроликов, завезенных в Австралию. Примером экспоненциального роста можно считать размножение микроорганизмов в загрязненных органическими и биогенными веществами водоемах.
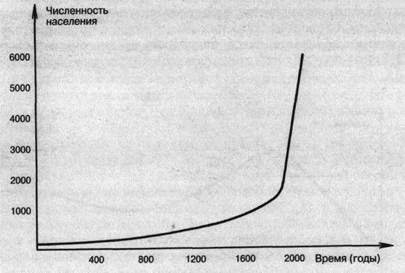
Рис. 14. Рост населения мира (млн чел.)
Близок к экспоненциальному типу рост населения популяции человека в настоящее время (рис. 15). Он обусловлен прежде всего резким снижением смертности в детском возрасте.
Естественный рост популяции никогда не реализуется в форме экспоненциальной кривой. В крайнем случае, если это и происходит, то в течение относительно короткого отрезка времени (как в приведенных выше примерах), после чего скорость роста численности снижается. Объясняется это тем, что не только в природных, но и в оптимальных экспериментальных условиях рост численности ограничен комплексом факторов внешней среды и реально складывается как результат соотношения меняющихся значений рождаемости и смертности.
Рост численности популяции замедляется по мере увеличения ее плотности, так как условия для роста и размножения особей становятся менее благоприятными. Например, животным при высокой плотности популяции может не хватать пищи. Растения начинают затенять друг друга, или им не хватает влаги. По мере ухудшения условий удельная скорость роста снижается, и при некоторой плотности численность популяции перестает расти. Эту предельную плотность, которой может достигнуть популяция в данных условиях, называют емкостью среды. Если рост популяции ограничен ресурсами, то после достижения емкости среды численность популяции колеблется возле некоторого среднего уровня и популяция подчиняется правилам логистического роста.
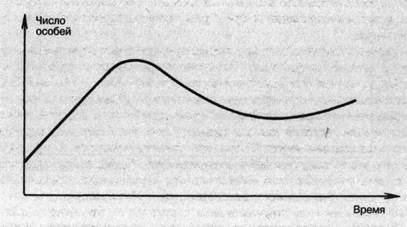
Рис. 15. Логистическая кривая роста популяции
График изменения численности популяции при логистическом росте представляет собой кривую, которая называется логистической кривой и напоминает по форме латинскую букву S (рис. 15). При логистическом росте популяции ее численность некоторое время нарастает, но вскоре этот процесс начинает замедляться, и постепенно рост численности практически прекращается. Для большинства популяций и видов выживаемость отображается логистической кривой.

