2015-10-13
2015-10-13 1067
1067Анализируя механизмы коммуникации у птиц, Е. Н. Панов (1978) приходит к выводу о «семантической полифункциональности» сигнала, который подчас столь всеобъемлющ, что приводит к полной его вырожденности. Типологическому подходу, рассматривающему коммуникацию как диалоговый обмен стереотипными дискретными сигналами, противопоставляется понимание коммуникации как длительного процесса, развертывающегося во времени в социально организованном пространстве. Кратко рассмотрим соотношение «дискретности» и «непрерывности» в акустической сигнальной системе птиц в гнездовой период. Примером крайне неспецифичного сигнала может служить песня самцов, которая используется как при «ключевых» социальных взаимодействиях (распределение территории, формирование пар, копуляция), так и в других ситуациях: отвлекающие демонстрации, контакт с птенцами, например у куликов. В то же время, ряд категорий сигналов (следования, тревоги при опасности на близком и дальнем расстоянии) обладают высокой специфичностью.
Функциональное значение сигнала может определяться взаимоотношением акустической и моторной подсистем коммуникации. Так, например, у кулика-воробья сигнал «преследования» нарушителя кормовой территории, излучаемый самцом—хозяином, в сочетании с определенными формами демонстративного поведения усиливается, а при других его формах теряет функциональный смысл. Некоторые категории сигналов имеют плавные взаимные переходы, например сигналы стайной тревоги и ее «отбоя» у кулика-воробья. Функциональную нагрузку в этом случае несут не только «полярные» категории этих сигналов, но и сам промежуточный вариант, отражающий успокоение стаи. Сигналы тревоги одиночной птицы и сигналы стайной тревоги характерны для совершенно аналогичной ситуации приближения потенциально опасного объекта. Однако динамическая организация этих двух сигналов различна: в стае сигналы излучаются с более высоким ритмом следования импульсов и более длительное время, что связано с взаимной стимуляцией птиц. Коммуникация у куликов при отвлекающих демонстрациях выражается в быстрой реакции «затаивания» птенцов в ответ на тревожные сигналы родителей и в согласованных действиях партнеров. Эффективность коммуникации в подобных ситуациях достигается по типу «экспресс-реакции» и не может рассматриваться как результат статистических процессов. Акустические сигналы, «запускающие» комплекс защитных реакций у куликов, голенастых, веслоногих, чайковых и воробьиных птиц, обладают высокой «специфичностью» не только в видовом, но и межвидовом плане. Адекватное соответствие сигналов и ответных экспресс-реакций при сбалансированном уровне мотиваций наблюдается и в других ситуациях, связанных с заботой о потомстве, у куриных и пластинчатоклювых, голенастых, чайковых и чистиковых и воробьиных птиц. Характер звучания сигнала в этих случаях может классифицировать биологическую ситуацию, а особенности его ритмической структуры могут отражать степень эмоционального возбуждения особи. У некоторых групп птиц, в частности у куриных, в процессе развертывания защитной реакции происходит «уточнение» ситуации, выражающееся в смене категорий сигналов, указывающих на характер опасности — появление наземного или пернатого хищника.
Успешное завершение гнездового периода имеет для популяции в целом (популяция, как известно, определяется в качестве элементарной эволюционной единицы) не менее важное значение, чем «ключевые» взаимодействия. Именно поэтому очевидна важная роль «специфичности» акустических средств в механизмах коммуникации у птиц. Высокая вариабельность сигнала может повышать, а не снижать эффективность коммуникации, иллюстрируя указанную закономерность увеличением разнообразия акустической сигнализации у высших позвоночных по сравнению с низшими (бесхвостыми амфибиями) и насекомыми.
Акустическая коммуникация у птиц может осуществляться как по «дискретному» принципу, так и обладать известной «непрерывностью» во времени. Подобный «двойственный» характер акустических сигнальных средств наряду с пластичностью поведенческих реакций обеспечивает необходимую адаптацию птиц к постоянно изменяющейся среде обитания и способствует поддержанию «популяционного гомеостаза».
Дискретность можно рассматривать как специализацию акустических сигнальных средств на осуществлении одной или нескольких функций (например, выделение сигналов тревоги, предупреждающих о появлении пернатого или наземного хищника, сигналов родительской опеки — пищевые сигналы, звуки сбора птенцов и т. п.). Однако подобная дифференциация приводит к увеличению числа дискретных сигнальных единиц, количество которых не может увеличиваться бесконечно. Именно поэтому коммуникация по «дискретному» принципу наблюдается преимущественно в тех случаях, когда существует необходимость обеспечения поведенческих «экспресс—реакций». При длительных взаимодействиях партнеров (брачные демонстрации и т. д.) обеспечивающих взаимную стимуляцию и синхронизацию физиологических состояний, отчетливо прослеживается «непрерывность» сигнальных средств, в том числе и акустических. Ранее мы подчеркивали, что наиболее значительным изменениям в процессе эволюции родственных групп птиц подвергаются сигналы, выполняющие при коммуникации «интегрирующие» и «синхронизирующие» функции. «Интегрирующие» и «синхронизирующие» эффекты достигаются при коллективной активации особей в выводке, группе или гнездовой колонии. Выделяются несколько типов акустической стимуляции и синхронизации поведения. В качестве первичного типа мы рассматриваем стимулирующие и синхронизирующие эффекты, свойственные птицам в эмбриональный период развития. Внешним проявлением подобных эффектов у выводковых является феномен синхронного вывода птенцов в кладке. Соотношение стимулирующего и синхронизирующего эффектов неравнозначно у различных групп птиц. Акселерация развития и синхронный вывод птенцов свойственны преимущественно страусообразным, куриным и пластинчатоклювым. Для птиц выводкового и полувыводкового типов развития, насиживающих кладку с первого отложенного яйца (гагары, поганки, кулики, чайковые), наиболее выражен эффект акселерации. Акустические взаимодействия можно рассматривать как один из факторов акселерации развития сенсорных и моторных механизмов в постнатальном онтогенезе птенцовых. Наблюдаемое ускорение метаболических процессов под действием акустической стимуляции позволяет предполагать, что активная «нагрузка» на анализаторные системы облегчает развитие и усиливает возбудимость нейронов, включенных в систему управления обменом.
Таким образом, акустические взаимодействия обеспечивают не только синхронизацию поведенческих реакций эмбрионов или птенцов, но и опосредованно сказываются на физиологических процессах.
В качестве второго типа мы выделяем интеграцию и синхронизацию сложных форм поведения (пищевого и оборонительного) у птенцов в выводках (семьях). Акустические взаимодействия птенцов, а также птенцов и взрослых птиц играют важную роль не только у выводковых и полувыводковых, но и у имматуронатных птиц. Они обеспечивают интегративную связь в выводках, их целостность, ориентацию в пространстве и осуществляются главным образом как «экспресс-реакции», не требующие длительных контактов во времени излучателя и приемника акустических сигналов.
Третий тип — координация и синхронизация поведения особей во временных стаях (в период кочевок, миграций и зимовок). Отличие, например, от взаимодействий второго типа состоит в резком увеличении количества потенциальных «излучателей» и «приемников», что создает повышенное дублирование сигналов и обеспечивает возможность опосредованной ориентации.
Длительные взаимодействия партнеров (например, в случае брачных демонстраций и подготовки к копуляции) обеспечивающие взаимную стимуляцию и синхронизацию их физиологического состояния, представляют собой четвертый тип. Сюда же относятся стимуляция и синхронизация некоторых физиологических состояний у птенцов в выводках, требующих длительных взаимодействий (например, переход от насыщения к отдыху и сну). Как правило, этот тип взаимодействий характеризуется разнообразием двигательных и акустических, реакций.
Множественная стимуляция и синхронизация физиологических состояний (от готовности к спариванию до яйцекладки и насиживания) — пятый тип акустических взаимодействий, характерный для колониальных птиц. Известно, что птицы, гнездящиеся в колониях, быстрее приступают к откладке яиц и размножаются более синхронно, чем особи того же вида, образующие одиночные гнезда. Данный тип взаимодействий складывался, по-видимому, параллельно с другими ключевыми признаками колониальности облигатного или факультативного типа. Возможность его формирования обусловлена эволюционной преемственностью адаптации, обеспечивающих популяционный. гомеостаз. Колония как биологическая система включает взаимодействия всех указанных выше уровней. Их соотношение в колониях птиц различных систематических и экологических групп имеет свою специфику. Именно поэтому мы выделяем особый, «колониальный» тип, представляющий собой комплексный механизм стимуляции и синхронизации физиологических и поведенчески реакций.
Принцип стимуляции «подобного подобным», к которому можно отнести описанные нами типы акустических взаимодействий является одним из основных механизмов, регулирующих поведение животных.
В заключение необходимо вернуться к рассмотрению причин редукции акустической системы коммуникации у целого ряд групп. Поскольку излучение звуковых сигналов у птиц сопряжено с функцией дыхательной системы, не подлежит сомнению такое ее свойство как мультифункциональность, проанализировано возникновение конфликтных ситуаций в мультифункциональных системах, ограничивающих возможность реализации гомеостаза в определенных условиях адаптации. На уровне органов в качестве примера подобного ограничения указано на невозможность одновременного участия мышц гомойотермных животных в процессах локомоции и терморегуляторной дрожи. Аналогичным образом у амфибий участие подъязычного аппарата в процессах принудительного вдоха и питания приводит к невозможности одновременного осуществления этих двух функций. Острота ограничения настолько велика, что в разных эволюционных ветвях амфибий либо редуцируются легкие, либо прикрепление языка переносится с подъязычного аппарата на нижнюю челюсть.
Примером подобного острого ограничения может служить, по-видимому, и редукция голоса у бескилевых птиц, в ызванная возрастным увеличением размеров трахеи (как длины, так и площади поперечного сечения), изменением пространственного расположения тимпанальных мембран сиринкса внутри бронхиальных проходов в процессе роста трахеи. Таким образом, этот случай наглядно иллюстрирует невозможность одновременного увеличения размеров трахеи при достижении видового морфобиологического облика и сохранения развитой вокализации. По-видимому, с аналогичным явлением связана значительная редукция голоса и у некоторых крупных видов голенастых птиц (колпицы, белого аиста и др.).
В конце XIX и в первой четверти прошлого века в России большой популярностью пользовалась так называемая "соловьиная охота". В те времена среди любителей природы было немало тонких знатоков соловьиного пения[37].
Когда натуралисту-птицелову удавалось поймать какого-нибудь выдающегося певца, этот соловей порой на несколько лет становился центром внимания всех поклонников маленькой серовато-бурой птички. Нюансы в пении каждого такого соловья служили в среде этих прекрасных знатоков русской природы предметом столь же детального и многостороннего обсуждения, какому подвергается среди меломанов исполнительское мастерство известного пианиста или скрипача. Существовала богатая и разнообразная терминология для обозначения колен соловьиного пения. Среди них различались, например, такие колена как "катушка", "гремушка"; стукотни -"перебивная", "дятловая", "юлиная"; свисты - "смирновский", "визговой", "польский", "подъемный"; дудки - "трелевая", "светлая", "водопойная", "лягушачья" и т. д.
В зависимости от того, какие из этих колен были наиболее характерны для прослушиваемого певца, его относили к одному из множества известных "сортов" - к "новосильским", "графским", "свистовым", "польским со свистовым ходом".
Известный русский натуралист И.К. Шамов, автор прекрасной, поэтической книги "Наши певчие птицы", так описывает впечатление, которое производило на слушателей пение одного из знаменитых московских соловьев: "Лучшая птица "графского" сорта, за все время, как он держался в охоте, была в 1874 г. у известного в то время охотника К.П. Смежевского; она была выставлена на публику в трактире "Прага" у Арбатских ворот и массу охотников собирала около себя своим выдающимся пением, стукотнями и дудками. И надобно было слышать, как исполняла птица эти песни... В жар и озноб бросало охотников, слушавших ее дудки и стукотни!"
Сегодня зоологи, исследующие поведение животных, располагают новейшей, совершенной аппаратурой, которая позволяет не только записывать звуки, но и переводить их в форму видимых изображений. И теперь мы можем попытаться достаточно точно и объективно оценить, как именно сочетается в песне соловья удивительное разнообразие звуков, радующее нас свежей новизной того или иного напева, с несомненной упорядоченностью мотивов, придающей импровизации нашего маленького певца качества подлинного музыкального произведения. В Западных предгорьях Алтая соловей в течение 40 минут исполнил 240 последовательных вариантов песни. Когда получили изображения всех этих песен и подсчитали число тех исходных звуков, из которых соловей компоновал свои мелодии, то оказалось, что на протяжении сорокаминутной записи их было 256. Можно предположить, что его возможности далеко не исчерпываются полученным перечнем исходных простейших звуков. Из 240 записанных песен только 11 имели своих абсолютных "двойников". Таким образом, в записи оказалось 229 различных песенных вариантов. Правда, среди них многие песни имели сходство с другими. Когда стали считать "одинаковыми" те песни, которые совпадали по числу и характеру фраз, но отличались по количеству нот в той или иной фразе, то стало возможным оценить число "разных" вариантов цифрой 122. Если же еще более огрубить степень сходства между отдельными песнями, рассматривая, например, состоящую из одной фразы песню в качестве незаконченной и "однотипной" с другими, начинающимися с такой же фразы, то число различных вариантов снизится до 74.
Большинство из этих 74 вариантов были спеты по 2 раза, но некоторые повторялись в записанной последовательности 8 и даже 11 раз. О чем это говорит? Мы видим, что разнообразие напевов соловья очень велико, но не беспредельно. Если бы птица компоновала исходные "ноты" совершенно случайным образом, то общее число доступных ей вариантов песен выражалось бы почти астрономической цифрой - 1049223 60! При таком числе вариантов вероятность повторной встречи хотя бы одного из них среди 240 песен была бы ничтожно мала. На самом же деле 58 из 74 вариантов встречены в записи более чем по одному разу, а некоторые - даже более 10 паз. Это значит, что соловей не только импровизирует, создавая по ходу своего пения новые комбинации известных ему нот, но и способен вновь и вновь возвращаться к уже готовым, "апробированным" вариантам, лишь немного разнообразя их звучание (например, меняя число нот в той или иной фразе или же добавляя к однажды спетому варианту одну, две или три фразы).
Оказывается, соловей обладает превосходной памятью. Он не только способен запомнить и абсолютно точно воспроизвести в любое время каждый из многих десятков песенных вариантов, включающих в себя до 15 и более исходных нот 5-6 разных типов. Соловей также держит в памяти и может повторить когда угодно целые группы таких сложнейших вокальных конструкций, включающие до 6 песенных вариантов. И это не просто механическое запоминание, поскольку при повторении подобных группировок из нескольких песен последние могут меняться местами. Например, сначала птица выстраивает различные напевы в порядке 1-2-3-4, потом воспроизводит еще 20 - 30 других песен, после чего вновь повторяет начальную группировку, но уже в ином порядке - скажем, 1 - 4 - 3 - 2 или же 1 - 3 - 2 - 4.
Создавая по ходу своей импровизации все новые и новые варианты напевов, соловей комбинирует исходные ноты не кое-как, а руководствуется при этом вполне определенными правилами" и "запретами". Он выбирает из своего обширного репертуара исходных звуков те или иные ноты с подходящими к случаю акустическими характеристиками.
Наиболее интересен в этом смысле "принцип контрастности' соседствующих фраз. Это значит, что, если, например, вторая фраза данной песни составлена из коротких высокочастотных нот, то следующая, третья фраза будет скорее всех содержать в себе продолжительные и низкочастотные звуки Кроме того, по характеру каждой ноты мы можем довольно определенно предсказать, будет ли она служить материалом для конструирования начальной, срединной или конечной фразы песни. В частности, ноты начальных фраз — это обычно продолжительные звуки без частотной амплитуды и с узким звуковым спектром, тогда как ноты конечных фраз, как правило, представляют собой короткие сигналы с широким звуковым спектром или с резкими перепадами частоты от начала к концу сигнала.
Здесь, правда, необходимо подчеркнуть, что все эти правила далеко не абсолютны и порождаемые ими запреты не слишком сильно ограничивают используемые соловьем возможности комбинирования нот.
Мы видим, что на первый взгляд организация песни соловья во многом напоминает структуру человеческой речи. И там и тут существуют исходные звуковые "кирпичики" (фонемы - в речи, ноты - в песне), из которых путем их комбинирования создаются достаточно длинные и сложные конструкции. Можно было бы пойти дальше и уподобить песенный вариант слову, а повторяемую группировку из нескольких вариантов - предложению. Поскольку число исходных нот в песне соловья весьма велико, а возможности их комбинирования ограничены лишь в небольшой степени, птица может создавать огромное количество дискретных "слов" и "предложений", запоминать их с абсолютной точностью и использовать в дальнейшем.
В этом смысле здесь есть внешняя аналогия с тем свойством нашего языка, которое называется открытостью, или продуктивностью. Благодаря этому качеству мы можем строить сколь угодно большое число разных высказываний из небольшого запаса элементарных звуковых единиц - фонем.
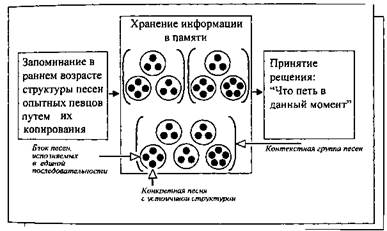
Рис.. Иерархический принцип хранения песенных вариантов в памяти соловья.
От 3 до 5 разных вариантов (черные фигурки) запоминаются как элементы компактных сочетаний (оконтурены окружностями). Несколько таких сочетаний птица держит в памяти совместно, в составе так называемых "контекстных групп" (большие скобки). Предполагается, что при пении соловей принимает ряд последовательных "решений", идя как бы от общего к частному: сначала он извлекает из памяти определенную контекстную группу, затем — сочетание песен и, наконец, некую конкретную песню.
Самец соловья, вернувшийся с места зимовки и занявший индивидуальный участок (территорию, как говорят зоологи), совершенно бессознательно извещает своим пением других самцов о том, что это место уже занято. Если бы соловьи обладали языком, они могли бы перевести бесконечную песню владельца участка так: "Здесь уже есть один самец, и Вам здесь делать нечего".
Когда ту же песню слышит вернувшаяся с зимовок соловьиха, она истолковывает чудесные трели примерно следующим образом: "Здесь уже есть самец. Почему бы не вступить с ним в законный брак?"
Но, как мы помним, самец соловья начинает петь еще во время весеннего пролета, когда он не готов ни к занятию территории, ни к приобретению подруги. В данном случае его пение свидетельствует лишь о физическом благополучии птицы и о том, что под влиянием удлиняющегося светового дня в ее крови накапливаются половые гормоны.
Можно задать вопрос: какие же преимущества дают соловью богатство и разнообразие его песен с их сложной внутренней структурой перед другими, гораздо менее искусными певцами? В то время как соловей на протяжении всей ночи так и эдак комбинирует сотни известных ему нот, создавая подлинные чудеса музыкальной импровизации, в соседних тростниках ему непрерывно вторит монотонное, бесконечное "зер-зер-зер-зер-зер...". Это поет обыкновенный сверчок - птичка чуть меньше соловья, столь же скромно окрашенная и мало чем отличающаяся от знаменитого певца по своему образу жизни. И все те потребности, которые удовлетворяются в общении соловьев с помощью сотен несходных нот и тысяч различных напевов, не менее успешно обслуживаются у речных сверчков одним единственным звуком "зер", повторяемым раз за разом с завидным постоянством.