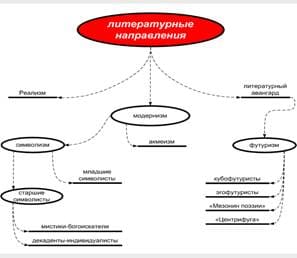
2013-12-31
2013-12-31 1535
1535ПОДКЛАСС ПОЛИПОДИЕВЫЕ, ИЛИ НАСТОЯЩИЕ ПАПОРОТНИКИ
Порядок Полиподиевые (Polypodiales)
Семейство схизейные (Schizaeaceae) - очень своеобразное древнее семейство, включающее 4 рода и 200 видов, обладает множеством примитивных признаков. Схизейные распространены в основном в тропических и субтропических областях на всех континентах. Большинство из них - растения средних размеров, имеющие короткие вертикальные или стелющиеся наземные побеги либо подземные корневища, от которых отходят перисторассеченные листья. Во влажных субтропических лесах встречаются очень мелкие виды схизеи (Schizaea) (рис. 61), анемии (Anemia) и актиностахиса (Actinostachys) размером 5-10 см. Их лентовидные или нитевидные листья не превышают 2 см и напоминают листья ситника. К этому семейству относится уже упоминавшийся род лигодиум, представляющий собой листовую лиану.
Чрезвычайно велико у схизейных разнообразие анатомического строения. У лигодиума на протяжении всей жизни сохраняется самый примитивный тип стелы - протостела. У схизеи и анемии протостела имеется лишь у молодого растения, а позднее формируется эктофлойная сифоностела, которая у схизеи сохраняется до конца жизни, а у анемии сменяется на диктиостелу. Таким образом, онтогенез стелы у схизейных отражает ход филогенеза стелы папоротников.
Семейство глейхениевые (Gleicheniaceae), известное еще с карбона, интересно сочетанием примитивных и высокоспециализированных признаков. Это одно из немногих семейств, большинство видов которого являются светолюбивыми ксерофитами. Они приурочены к засушливым областям субтропических и умеренных областей в основном Южного полушария. Большинство из них имеют длинные корневища, от которых отходят листья либо прямостоячие олиственные побеги. Листья довольно крупные, у некоторых представителей длиной 1,5-2 м и более. Наиболее крупный род глейхения (Gleichenia) имеет жесткие листья с узкими долями чрезвычайно своеобразного облика. Верхушка его листа довольно быстро заканчивает свой рост благодаря заложению острой спящей почки, часто имеющей острую жесткую вершину. Продолжение роста листа происходит за счет боковых перьев, которые также заканчиваются почкой и т.д., в результате лист приобретает объемное ложнодихотомическое ветвление.
Большой древностью семейства объясняется примитивный тип стелы - у одних видов сохраняется протостела, у других она сменяется амфифлоической сифоностелой. Диктиостела у глейхениевых отсутствует.
Семейство гименофилловые (Hymenophyllaceae) - одно из удивительных и интереснейших семейств папоротников, поражающих красотой своих форм и нежностью прозрачных листьев (гименофилловые переводятся как пленчатолистные). Гименофилловые - мелкие папоротники, живущие во влажных тропических и субтропических лесах на почве, эпифитно на мелких мхах, селягинеллах, на основании стволов деревьев, где постоянно влажно и темно. Часто определенный вид имеет свою строго определенную экологическую нишу. Приспособительная эволюция гименофилловых пошла по пути резкой редукции спорофита, что и придает своеобразие этой группе. Лишь у немногих представителей листья достигают 50-60 см, у большинства же они не превышают 2-3 см, а у некоторых видов - 3-4 мм. Уникальность листьев гименофилловых проявляется в том, что у большинства видов листовые пластинки однослойные, реже 2-4-слойные, поэтому они прозрачные и почти бесцветные (см. рис. 61). У большинства видов листья перисторассеченные, реже цельные, двумя рядами располагаются на тонких, длинных корневищах; от них же отходят тончайшие корни. В случаях крайней редукции функцию корней выполняют многочисленные волоски на всех частях растения. Листья, лишенные эпидермы, при относительной влажности ниже 90 % страдают от иссушения, и растение в этом случае переходит к состоянию анабиоза. Мелкие наземные гименофилловые образуют сплошной покров на поверхности земли, создавая влажную микросреду. У скальных видов листья густо покрыты волосками и способны в сухие периоды скручиваться в трубочки. Сорусы располагаются либо на зеленых листьях, либо на специализированных спорофиллах, по краю листа, либо на вершинах перышек. Сорусы окружены своеобразным покрывальцем, каждый сорус снабжен жилкой.
Семейство циатейные (Cyatheaceae) содержит около 1000 видов, приуроченных в основном к тропическим и субтропическим областям. Это древнее семейство объединяет крупные древовидные формы, переходные к травянистым, и типичные травянистые длиннокорневищные формы. Семейство характеризуется как примитивными, так и специализированными чертами. Проводящая система у разных видов варьирует от протостелы до простой и полициклической диктиостелы. Сорусы у одних видов занимают краевое положение, у других они расположены на нижней стороне вдоль жилок. Сорусы прикрыты специализированными или малоспециализированными покрывальцами. Спорангии крупные, с косым кольцом, расположены, как правило, на удлиненной плаценте.
Древовидные циатеи и альзофилы приурочены в основном к влажным прохладным горным областям тропиков и субтропиков. Наибольшее число видов (около 600) содержит род циатея (Cyathea). Стволы, достигающие у некоторых видов 20 м высоты, несут крупные, до 6 м длины, дважды- или триждыперистые листья с длинными черешками. К циатее близок род диксония (Dicksonia), содержащий 25 видов и распространенный в Австралии с прилегающими островами и в Южной Америке. Они произрастают в горных лесах. Большинство видов представлено древовидными формами высотой до 4-6 м или переходными полудревовидными с короткими толстыми стеблями. Остальные циатейные представлены либо переходными формами с коротким стволом, либо типичными длиннокорневищными растениями. К ним относится монотипный род орляк (Pteridium aquilinum), распространенный по всему земному шару. Орляк образует обширные заросли благодаря обильно ветвящемуся корневищу (рис. 62). Во многих странах он находит применение как пищевой продукт. Остальные корневищные папоротники этого семейства относятся в основном к тропическим видам.
Семейство асплениевые (Aspleniaceae) - наиболее молодое, самое многочисленное (до 400 видов) и наиболее распространенное семейство как в тропических, так и в умеренных областях. Оно содержит много эволюционно продвинутых признаков, характеризуется обилием жизненных форм, отражающих экологические условия их обитания, и разнообразием анатомо-морфологических структур листьев и стеблей. Преобладают длинно- и короткокорневищные папоротники различного облика, реже встречаются виды с прямостоячими побегами; лиан и эпифитов очень мало. Лишь несколько видов рода блехнум (Blechnum) имеют древовидную форму высотой 1-1,5 м.
Наиболее многочисленный род асплениум, или костенец (Asplenium), объединяет около 700 видов, распространенных по всему земному шару. Виды умеренных областей имеют размеры в несколько сантиметров, у тропических папоротников листья достигают в длину 1,5-2 м; многие виды размножаются листовыми почками. Некоторые тропические виды представлены эпифитами, среди них наиболее эффектен описанный выше папоротник асплениум птичий (A. nidus). К роду асплениум близок род листовик (Phyllitis), один из видов которого - " олений язык ", или " тещин язык " (Ph. scolopendrium), распространен в лесах Кавказа, Европы, Юго-Восточной Азии, Северной Америки. Он имеет жесткие удлиненно-овальные листья длиной до 60 см, похожие на листья цветковых растений и отличающиеся от них только по наличию линейных сорусов на их нижней стороне.
Крупный род щитовник (Dryopteris) (см. рис. 62) содержит 150 видов, которые в основном произрастают в умеренных областях Северного полушария. Наиболее широкий ареал имеет щитовник мужской (D. filix-mas), обладающий коротким корневищем и розеткой дважды перисторассеченных листьев. Он произрастает в лесах Европы, Азии, Америки, на скалах Гренландии и Скандинавии. Близкий к нему род кочедыжник (Athirium) содержит около 200 видов, приуроченных также к умеренным областям Северного полушария. Наиболее распространен кочедыжник женский (A. filix femina), отличающийся большой рассеченностью нежной листовой пластинки. Подобный же ареал имеет голокучник (Gymnocarpium) с наиболее распространенным видом голокучником трехраздельным, или папоротником Линнея (G. dryopteris). Этот небольшой длиннокорневищный изящный папоротник весьма загадочно выглядит весной, когда каждая его доля свернута в тонкую улитку, а сам он напоминает доисторическое растение в миниатюре. Самым величественным из числа северных папоротников можно считать страусник (Matteuccia), который на влажных, относительно богатых почвах формирует изысканные воронки высотой до 1-1,5 м из перистых листьев. Они имеют красивую листовую пластинку низбегающей формы, напоминающую перо страуса. Ранней весной листья покрыты голубоватым восковым налетом, и воронки кажутся светящимися изнутри. Летом появляются невзрачные светло-зеленые спорофиллы, буреющие к осени.
21-24 ОТДЕЛ
ГОЛОСЕМЕННЫЕ (PINOPHYTA, ИЛИ GYMNOSPERMAE)
Голосеменные - древняя группа, берущая свое начало с верхнего девона и органично связанная о группой праголосеменных растений. Расцвет голосеменных приходится на мезозойскую эру, когда господствовали теплолюбивые виды. К концу мезозоя происходит сокращение таких теплолюбивых форм, как беннеттитовые, саговниковые, семенные папоротники. Второе рождение голосеменные обрели благодаря появлению холодостойких видов в недрах мезозойской флоры. Возможно, что их возникновение было связано с местными внутриматериковыми оледенениями. Эти виды оказались способными в кайнозойскую эру осваивать обширные пространства в умеренных и умереннохолодных областях Северного полушария, где они и поныне на больших территориях занимают господствующее положение в сложении растительного покрова. Широкое распространение голосеменных в значительной степени объясняется тем, что размножение у них осуществляется семенами. Семя как зачаток нового организма представляет собой комплексный многоклеточный орган, состоящий из зародыша, запасающих тканей эндосперма и многослойной семенной кожуры, надежно обеспечивающей сохранность зародыша. У большинства голосеменных семена обладают периодом покоя. Таким образом, молодые растения вступают в самостоятельную жизнь более жизнеспособными, чем споровые растения.
При прорастании семени первым появляется зародышевый корешок, который формирует стержневую корневую систему; лишь у немногих видов главный корень заменяется системой боковых или придаточных корней. Наличие главного корня явилось базой для возможного формирования крупных древесных форм. Этому способствовало также наличие длительно функционирующего камбия, который формирует древесину - водопроводящую ткань, обладающую большой механической прочностью. Среди современных голосеменных преобладают древесные формы, однако важно подчеркнуть, что среди древних палеозойских видов в разных систематических группах голосеменных встречались формы травянистые, кустарниковые, древесные со слабо развитой древесиной. Среди современных саговниковых имеются древесные формы со значительно развитой сердцевиной и корой, виды с клубневидными побегами полудревесной и типично травянистой жизненных форм. У подавляющего большинства голосеменных побеги нарастают моноподиально. Примитивные голосеменные характеризуются отсутствием ветвления, большинство же ныне живущих видов обильно ветвится. Листья всех голосеменных, несмотря на колоссальное разнообразие их форм и размеров, имеют синтеломное происхождение. Из числа современных древние виды имеют крупные, сильно рассеченные листья, у большинства же видов листья мелкие, игловидные или чешуевидные. Древесина голосеменных состоит в основном из трахеид, паренхима развита довольно слабо, либриформ отсутствует. Характерным признаком трахеид голосеменных является наличие окаймленных пор, ровными рядами расположенных на радиальных и тангентальных стенках. Для флоэмы характерно отсутствие клеток-спутниц.
Все голосеменные - разноспоровые растения; в подавляющем большинстве случаев имеются микро- и мегастробилы различной формы, которые, в свою очередь, могут объединяться в констробилы. В микроспорангиях формируются микроспоры, которые начинают прорастать под оболочкой. Проросшие микроспоры - мужские гаметофиты, заключенные в оболочку микроспоры, называются пылинками, а совокупность пылинок - пыльцой. После первого деления микроспоры возникает одна маленькая клетка и вторая - более крупная, которая может отчленить еще одну маленькую клетку (рис. 66). Эти первые (одна или две) клетки соответствуют вегетативной части гаметофита (по-гречески prothallium), поэтому называются проталлиальными. Более крупная клетка, получившая название антеридиальной инициали, делится еще раз с образованием двух очень специализированных клеток. Одна из них формирует антеридий и поэтому называется антеридиальной. Судьба другой клетки зависит от типа полового процесса. У наиболее примитивных групп, у которых половой процесс осуществляется с помощью сперматозоидов, из второй клетки развивается гаустория, поэтому ее называют гаусториальной. У высших голосеменных, у которых половой процесс происходит с помощью спермиев, из этой клетки развивается пыльцевая трубка, поэтому ее называют клеткой трубки, или сифоногенной. Часто ее называют вегетативной, хотя к вегетативной части гаметофита она не имеет никакого отношения. Антеридиальная клетка делится на 2 клетки, одна из которых либо образует 2 сперматозоида, и тогда называется сперматогенной, либо 2 спермия, и тогда называется спермагенной. Вторая клетка антеридия остается стерильной и впоследствии разрушается. Разные авторы, по-разному ее интерпретируя, называют эту клетку ножкой, стенкой антеридия, дислокатором, сестринской клеткой, внося терминологическую неясность; поэтому предложено называть ее просто стерильной клеткой. Пылинка, состоящая у различных видов из разного количества клеток, разносится ветром. Поскольку у голосеменных в отличие от покрытосеменных нет замкнутой завязи, пылинка попадает непосредственно на семязачаток.
Развитие семязачатка начинается с появлением срединного бугорка - нуцеллуса, внутри которого на ранних стадиях развития выделяется одна крупная спорогенная клетка (рис. 67). Позднее она подвергается редукционному (мейотическому) делению и образует 4 неравноценные клетки - мегаспоры. Поэтому нуцеллус следует рассматривать как мегаспорангий. Вслед за бугорком - нуцеллусом из периферической меристемы возникает 2 бугорка, которые, разрастаясь, начинают формировать валик. Он обгоняет в своем росте нуцеллус, нависая над ним и образуя интегумент с небольшим отверстием - микропиле, или пыльцевходом. Таким образом, семязачаток представляет собой сложный орган, состоящий из мегаспорангия - нуцеллуса и покрова - интегумента. По поводу происхождения интегумента существует ряд гипотез. Согласно одной из них, синангиальной, семязачаток возник из синангия, у которого внутренний спорангий остался фертильным (т.е. продуцирует споры), а периферические подверглись стерилизации и образовали интегумент. Согласно другой, теломной, теории интегумент представляет собой результат срастания периферических изначально вегетативных теломов вокруг одного спороносного. Эта теория согласуется с палеоботаническими находками, которые относятся к праголосеменным и вымершим голосеменным - семенным папоротникам.
Из четырех образовавшихся мегаспор 3 мелкие погибают, а крупная прорастает в женский гаметофит, или заросток, который у голосеменных называется эндоспермом. После оплодотворения из семязачатка формируется семя. Интегумент превращается в семенную кожуру, нуцеллус расходуется на развивающийся зародыш, от него остается тонкая пленочка. Ткани заростка или эндосперма сильно разрастаются, и в них откладываются запасные питательные вещества. Из оплодотворенной яйцеклетки формируется зародыш, состоящий из корешка, стебелька и почечки с 2-18 семядолями.
Классификация голосеменных основана на разных подходах. Согласно новейшей классификации, отдел объединяет 6 классов; из них 2 класса вымерших растений - семенные папоротники и беннеттитовые и 4 класса ныне живущих - саговниковые, гинкговые, оболочкосеменные и хвойные.
КЛАСС СЕМЕННЫЕ ПАПОРОТНИКИ (PTERIDOSPERMOPSIDA, ИЛИ LYGINOPTERIDOPSIDA)
Это самая древняя, полностью вымершая группа голосеменных, жившая с середины девона до мелового периода; расцвет их приходился на каменноугольный период. В эту группу объединяют растения, имевшие папоротниковый облик, но формировавшие семязачатки. Эти древние растения представляют большой теоретический интерес, так как, с одной стороны, вскрывают разнообразие анатомо-морфологических структур, отражающих возможные пути их эволюции, а с другой стороны, демонстрируют различные варианты формирования семязачатков, разнообразие их форм и структур. По сохранившимся остаткам трудно реконструировать облик этих растений; описаны фрагменты растений с диаметром стволов 1,5-2 м и 20-30 см. Некоторые из них имели форму кустарников, мелких трав с диаметром стеблей от 2-3 см до 2-3 мм. Одни из них не ветвились, другим было свойственно дихотомическое или боковое ветвление; описаны даже укороченные побеги. На стеблях некоторых растений имелись воздушные корни. Многообразие жизненных форм сочеталось с различными вариантами строения и расположения листьев. Листорасположение было очередное и мутовчатое, листья располагались по всему стеблю или только в его верхней части. Листья были голые, покрытые железами или волосками, они различались по характеру и степени расчленения (перистые и пальчатые, однажды- и многократно рассеченные, цельные), по форме перышек (овальные, яйцевидные, линейные), по характеру жилкования (открытое, сетчатое).
Не менее разнообразно было анатомическое строение стеблей семенных папоротников. У одних видов камбий функционировал очень слабо, откладывая лишь несколько слоев вторичной ксилемы (травянистый тип стебля), у других растений имелась мощная древесина типа хвойных, у третьих - большую часть стебля составляла паренхима сердцевины, как у саговниковых. Подобное разнообразие морфологического и анатомического строения, которое обнаруживалось у разных растений в пределах одного периода их жизни, позволяет предположить, что формирование жизненных форм и анатомических структур как голосеменных, так и покрытосеменных с самого начала могло пойти разными путями, т.е. могли возникать первично древесные и первично травянистые формы. Поэтому из этой группы семенных папоротников легко вывести все прочие классы голосеменных растений.
Большой интерес представляет многообразие в строении семязачатков, которое проливает свет на происхождение интегумента. Самые древние семязачатки, описанные из верхнего девона и нижнего карбона (Archaeosperma, Hydrasperma, Genomosperma, Lagenostoma) (рис. 68), имели весьма примитивное строение - нуцеллус (мегаспорангий) с оттянутым на вершине носиком был окружен узкими, отогнутыми наружу лопастями интегумента. Лопасти срастались между собой и с нуцеллусом лишь в самом основании; в каждую лопасть входил проводящий пучок. Этот лопастный интегумент очень напоминал совокупность сближенных и начавших срастаться вегетативных теломов, что подкрепляет позиции синтеломной теории происхождения интегумента. У большинства семязачатков, описанных из среднего и верхнего карбона, лопасти интегументов полностью или почти полностью срастались между собой, оставаясь свободными лишь на вершине. Параллельно этому происходило срастание интегумента с нуцеллусом. У многих семенных папоротников одиночные семязачатки или группы семязачатков располагались в окружении чашевидного образования - купулы. Одни авторы рассматривают ее как выросты листа (по наличию железок на ее поверхности), другие считают ее результатом срастания периферических теломов. Купула описанных выше самых древних семязачатков представляла собой совокупность дважды или трижды дихотомически ветвящихся лопастей, в каждую из которых входила жилка. Наподобие лопастей интегумента они сливались лишь у основания. У среднекарбоновых и верхнекарбоновых представителей лопасти купулы, снабженные проводящими пучками, срастались либо на большем своем протяжении (Lagenostoma, Calathospermum), либо полностью (Mitrospermum). Купулы, в свою очередь, могли срастаться в большей или меньшей степени с интегументом семязачатков, формируя внешний интегумент (купуломент), пронизанный пучками (рис. 69). Подобный двойной интегумент имеется у семязачатков саговниковых. У более поздних голосеменных могло происходить полное слияние внутреннего интегумента с нуцеллусом вплоть до исчезновения проводящих пучков. В этом случае семязачатки с одним внешним интегументом становились вторично однопокровными. Таким образом, первые голосеменные могли иметь 3 варианта строения семязачатков - исходно однопокровные, т.е. с одним интегументом, двупокровные - с внутренним и внешним интегументами и вторично однопокровные с внешним интегументом. Кроме того, в пределах этих трех вариантов семязачатки имели радиальную или дорзивентальную симметрию, т.е. число вариантов строения семязачатков увеличивалось до 6. Помимо этого имелись еще дополнительные варианты семязачатков. Строение семязачатков оказалось очень стабильным и характерным для разных классов голосеменных растений. Многообразие семязачатков позволило установить преемственность репродуктивных органов покрытосеменных от голосеменных растений.
Семязачатки у одних видов располагались на обычных листьях, у других - на специализированных листьях - мегаспорофиллах, у третьих - на осевых структурах - мегаспорангиофорах. Интересная особенность семян семенных папоротников заключалась в отсутствии зародышей; по-видимому, семена опадали раньше, чем происходило полное их формирование, как у саговников. Микроспорангии часто объединялись в синангии, которые, как правило, располагались на специализированных перышках вегетативных листьев, т.е. имел место диморфизм частей листа. Форма спороносных частей листа была самая разнообразная. В микроспорангиях некоторых видов найдены мужские гаметофиты; у одних видов число проталлиальных клеток доходило до 30, а у некоторых имелось всего лишь 2 клетки.
КЛАСС ГИНКГОВЫЕ (GINKGOOPSIDA)
Класс гинкговые содержит одно семейство Ginkgoaceae с одним современным видом гинкго двулопастное (Ginkgo biloba). Это очень своеобразный класс, в котором сочетаются такие примитивные признаки, как оплодотворение с помощью сперматозоидов с очень совершенным типом ветвления и формированием мощной древесины, которая обеспечивает оптимальное водоснабжение и большую прочность ствола.
Представители гинкговых известны с конца каменноугольного периода, но расцвет их приходился на триас и юру, когда их многочисленные представители были широко распространены по всему земному шару. Из мезозоя описаны гинкговые, относимые к 8 порядкам (пельтаспермовые, каламопитиевые, каллистофитовые, гинкговые, лептостробовые, или челаковские, кейтониевые, арбериевые, или глоссоптерисовые, пентаксилеевые). Они были представлены разнообразными жизненными формами; описаны крупные деревья, мелкие кустарники с диаметром стволов до 3 см, травянистые виды и даже лианы. Примечательно, что у некоторых раннемеловых гинкговых наряду с обычными побегами формировались укороченные побеги. Большинство видов описано по листьям, стеблям либо по репродуктивным органам. Для стеблей большинства гинкговых было характерно наличие секреторных каналов. Вторичная древесина была сложена трахеидами с точечными окаймленными порами и пронизана сердцевинными лучами. Наибольшего разнообразия достигали листья гинкговых; прослеживается эволюционная тенденция перехода от многократно дихотомически рассеченных листьев через дихотомически лопастные к цельным.
В основу деления на порядки положено многообразие форм микро- и мегаспорангиев и характер их объединения. Микроспорангии у наиболее примитивных видов располагались на отдельных перышках вегетативных листьев, а у более продвинутых видов - на специализированных разветвленных спорангиофорах. Часто они объединялись в гроздевидные собрания, реже спорангии срастались в синангии. Семязачатки у палеозойских видов располагались одиночно на обычных листьях, а у мезозойских видов - на особых специализированных органах различной формы - перистой, щитковидной, купуловидной, либо на коротких перышках перистых спорофиллов. Намечалась тенденция к сокращению семязачатков до 1-2, параллельно этому у некоторых видов края купулы смыкались друг с другом, образуя почти замкнутую полость, внешне сходную с завязью покрытосеменных. Семязачатки всех гинкговых обладали двусторонней симметрией. Их единственный интегумент у одних видов был свободный, а у других срастался с нуцеллусом.
Род гинкго появился с верхнего триаса, в юре он был широко распространен в Европе, Азии, Северной Америке, Австралии и даже в Арктике. У мезозойских видов гинкго четко прослеживается переход от листьев многократно дихотомически рассеченных до лопастных и цельных. Характерной особенностью листьев ныне живущих и вымерших видов является вхождение в черешок двух жилок, каждая из которых многократно дихотомически ветвится. Разветвления этих жилок не анастомозируют между собой. У некоторых юрских видов обнаружены укороченные побеги с пучком листьев.
Современный вид - гинкго двулопастное - с давних времен считается священным храмовым растением, оно сохранилось благодаря защите его человеком. Обнаруженная в горах Китая небольшая рощица из гинкго представляет собой сохранившиеся остатки то ли естественных лесов, то ли культурных посадок. Гинкго широко культивируется в Японии, Корее, Европе и Северной Америке. Гинкго - это листопадное дерево высотой до 30 м; его красивая крона образована побегами двух типов - обычными удлиненными - ауксибластами и многочисленными боковыми - брахибластами длиной 2- 3 см. Обильное ветвление в сочетании с листопадностью приводит к ежегодному обогащению растения многочисленными физиологически обновленными листьями, что обеспечивает высокую интенсивность процесса фотосинтеза. Развитие укороченных побегов создает возможность наилучшего освещения всей листовой поверхности за счет эффекта листовой мозаики. Листья имеют длинный черешок и своеобразную веерную листовую пластинку. На удлиненных побегах образуются листья двулопастные и двураздельные, а на укороченных - цельные или слегка лопастные, с волнистым краем. В стволах гинкго, в отличие от саговников, сердцевина развита слабо. Большая часть ствола занята мощной древесиной, сложенной точечными трахеидами. Важной чертой трахеид гинкго является размещение пор на тангентальных стенках, что обеспечивает наилучшее снабжение водой камбия и способствует его активной деятельности.
В стадию спороношения гинкго вступает на 25-30-м году жизни. Микро- и мегастробилы, формирующиеся в конце лета на укороченных побегах в пазухах листьев, развиваются на разных растениях. Пыление происходит ранней весной до распускания листьев. Наилучшему рассеиванию пыльцы способствует форма микростробила в виде поникающей и раскачивающейся сережки. На длинной оси микростробила спирально располагаются микроспорангиофоры в виде тонкой ножки, на конце которой висят 2 микроспорангия. Вылетающая пылинка состоит из двух проталлиальных клеток - антеридиальной и гаусториальной. Мегастробилы гинкго возникают на укороченных побегах в количестве 5-7 и имеют необычную форму. Они напоминают веточку дуба с желудями. Ось мегастробила в виде длинной ножки заканчивается на вершине двумя семязачатками с расширенным валиком у основания. В оси проходят 2 пары пучков, каждая из которых подходит к семязачатку. Изредка ось на вершине разветвляется на 5-7 веточек, в этом случае в ось стробила входят 5-7 пар пучков. Следовательно, веточки, несущие семязачатки, представляют собой мегаспорангиофоры. Обычно у гинкго они редуцированы практически полностью, и семязачатки непосредственно сидят на оси мегастробила. Природа валика, окружающего семязачаток, неизвестна и дискутируется до сего времени. Семязачаток гинкго по своему строению напоминает семязачаток саговника и отличается от него лишь деталями. Развитие гаустории, процесс оплодотворения, формирование зародыша и семени в целом также похожи на саговниковые. Если пыление происходит весной, то оплодотворение осуществляется только осенью, иногда в уже опавших семязачатках, ни внешне, ни по размерам не отличающихся от созревших семян. Они имеют внешний мясистый слой семенной кожуры, срединный каменистый (склеротеста) и внутренний, похожий на пергамент. Семена прорастают без видимого периода покоя, что относится к числу примитивных признаков. Семена гинкго используются в медицине и употребляются в пищу. Будучи весьма декоративным растением, гинкго культивируется для озеленения южных городов.
Порядок Гнетовые (Gnetales)
Подобно вельвичиевым этот порядок содержит одно семейство Gnetaceae с одним родом гнетум (Gnetum), объединяющим 30 видов (рис. 75). Гнетумы распространены во влажных тропических лесах Азии, Южной Америки и Западной Африки, причем для каждого материка характерны свои эндемы. Большинство видов гнетума - лианы, но встречаются небольшие деревья и кустарники. Все виды гнетума имеют широкие, цельные, супротивно расположенные листья с сетчатым жилкованием, они похожи на листья двудольных. У некоторых видов по краям листьев образуются выводковые почки. Все виды гнетума также двудомные; их сережковидные констробилы построены по типу констробилов эфедры, но мутовки чешуевидных листьев спаяны у основания в кольцо. Микростробил чрезвычайно простой; он состоит из узкого, длинного трубчатого околоцветника, окружающего тычинку. Мегастробил сведен до одного семязачатка, который находится в окружении 2 пар сильно видоизмененных прицветных листьев. Семязачатки покрыты двумя интегументами: наружный - короткий, внутренний наверху вытянут в микропилярную трубочку. Оба интегумента срастаются снаружи с прицветными листьями, а внутри - с основанием нуцеллуса. Одна из 4 мегаспор прорастает в сильно редуцированный женский гаметофит, отдаленно напоминающий зародышевый мешок покрытосеменных. Он не имеет тканевой структуры и лишен архегониев.
Женский гаметофит имеет удлиненную форму, расширенную наверху и суженную внизу. На первых этапах его развития под оболочкой мегаспоры образуются многочисленные ядра, расположенные в постенном слое цитоплазмы. Затем в нижней части между ядрами возникают перегородки, и эта часть гаметофита становится многоклеточной; верхняя часть остается неклеточной.
Мужской гаметофит еще более редуцирован, чем у эфедры, так как он не имеет проталлиальных клеток. В цитоплазме пыльцевой трубки содержится лишь 3 ядра - самой пыльцовой трубки и 2 спермиев. Важно отметить, что оба спермия могут сливаться с ближайшими ядрами женского гаметофита, что напоминает процесс так называемого двойного оплодотворения покрытосеменных. Полного развития достигает лишь один зародыш.
При созревании семян внешний интегумет образует сочный мясистый слой ярко-розового цвета. Внутренний покров становится твердым, каменистым. Своеобразие в строении стробилов представителей класса гнетовых дает основание некоторым авторам связывать их в своем происхождении с беннеттитовыми.
Сходство строения женского и мужского гаметофитов, гнетовых с гаметофитами покрытосеменных дает основание некоторым эмбриологам сделать заключение о том, что строение зародышевого мешка и двойное оплодотворение покрытосеменных не носит характера уникальности.
ПОДКЛАСС ХВОЙНЫЕ (PINIDAE)
Этот подкласс наиболее многочисленный из всех голосеменных растений (он включает 7 семейств, 55 родов и около 600 видов), наиболее широко распространен практически по всему земному шару, кроме Арктики и Антарктиды. В истории Земли хвойные появились во второй половине палеозоя - из верхнего карбона описаны представители семейства лебахиевых (Lebachiaceae). Они зародились в Северном полушарии, по-видимому, в условиях умеренного климата; есть предположение, что локальное материковое оледенение вызывало увеличение сухости климата. Поэтому первые хвойные обладали чертами ксероморфной структуры и характеризовались наличием годичных колец в древесине. На границе перми и триаса произошло увеличение континентальности климата, поэтому в Северном полушарии резко возросло количество хвойных. Появляются наиболее древние из ныне живущих - араукариевые, подокарповые и сосновые. Однако четкого разграничения на семейства еще не было; так, виды араукарии, описанные из юры, сочетали в себе признаки сосновых, кипарисовых и араукариевых. В конце перми хвойные начали проникать в Южное полушарие, а с триаса происходит их широкое распространение по земному шару. Расцвета хвойные достигли в юрском и меловом периодах; к этому времени относится появление всех остальных ныне живущих семейств. В начале кайнозойской эры начинается обособление климатических зон и формирование ареалов растений. В первой половине третичного периода - в эоцене климат был еще теплый и влажный и хвойные были распространены от Арктики (они отмечены на Шпицбергене, в Гренландии, в Шотландии) до Антарктиды. В последующем периоде - в олигоцене ареал хвойных начинает сокращаться, но на территории Европы, Северной Америки, Восточной Азии произрастали сосновые леса с большим участием туи, пальм, каштанов, магнолий и других теплолюбивых видов. К олигоценовым отложениям относятся знаменитые залежи янтаря; бурые угли Германии образованы болотным кипарисом. Он в то время вместе с секвойей еще доходил до Шпицбергена, а в настоящее время произрастает в основном в устье реки Миссисипи. Хвойные в олигоцене встречались и в Антарктиде. Переломным периодом в истории хвойных оказался четвертичный период с серией оледенений. В Европе и Азии при наступлении ледника все теплолюбивые виды хвойных вымерли. Сохранились лишь молодые холодоустойчивые виды, которые после отступления ледника широко распространились в пределах умеренных и умеренно холодных областей. Исключение составила Юго-Восточная Азия, которая не подвергалась оледенению и сохранила много древних теплолюбивых видов. В Северной Америке горы расположены в основном в меридиональном направлении, поэтому теплолюбивые виды во время оледенения мигрировали на юг. После таяния ледника некоторые из них частично вернули свои позиции, хотя ведущая роль в настоящее время и в Северной Америке принадлежит молодым холодоустойчивым видам. В Южном полушарии сохранилось много древних видов хвойных, но они распространены в основном в областях с умеренным климатом. В тропиках хвойные встречаются главным образом в горах, т.е. также в условиях умеренного климата, хотя отдельные виды сосны и подокарпуса произрастают в субтропических лесах.
Современные хвойные представлены в основном деревьями, реже кустарниками или особой стланниковой - стелющейся формой. Однако среди древних вымерших хвойных встречались даже травянистые растения. Побеги у хвойных либо только удлиненные, либо двух типов - удлиненные и укороченные. Несмотря на название, листья хвойных далеко не всегда имеют форму хвои. Встречаются хвойные с яйцевидной, ланцетной формой листа, но чаще они имеют игловидную или чешуевидную форму. Судя по скудным палеоботаническим данным пермских лебахиевых, можно заключить, что они имеют синтеломное происхождение, так как листья их дихотомически ветвились или были на вершине раздвоены. Соответственно этому дихотомически ветвились и их жилки. Подавляющее число хвойных - вечнозеленые растения; продолжительность жизни листьев определяется как биологическими особенностями пород, так и экологическими условиями их жизни. У светолюбивых пород листья живут от 2 до 5 лет, у теневыносливых до 10-15 лет, а в высокогорных условиях - даже до 25-30 лет. Все вечнозеленые породы имеют жесткие, сильно кутинизированные и склерофицированные листья; устьица у них располагаются на нижней стороне в углублениях, что значительно сокращает испарение воды в зимний период. У листопадных пород - у лиственницы (Larix) и лжелиственницы (Pseudolarix) хвоя нежная, мягкая, как у обычных листьев, за что они и получили свое название. Для некоторых хвойных, например для таксодиума (Taxodium), метасеквойи (Metasequoia), сосны (Pinus), характерен веткопад, при котором опадают не отдельные листья, а целые укороченные веточки.
Корневая система, как правило, стержневая, но у некоторых видов, например у ели, главный корень довольно быстро замирает и формируется поверхностная корневая система. В других случаях, как, например, у сосны, корневая система оказывается довольно лабильной и ее характер определяется уровнем грунтовых вод. Для большинства хвойных характерны различные типы микоризы. Вегетативное размножение большинству хвойных не свойственно, исключение составляют пихта и секвойя, способные образовывать поросль. При обработке ауксинами некоторые хвойные приобретают способность вегетативно размножаться (таким путем размножают голубые ели).
Анатомическое строение стеблей хвойных характеризуется слабым развитием паренхимы сердцевины и мощным развитием древесины. Поры у разных видов могут располагаться как на радиальных, так и на тангентальных стенках. Отличительной чертой в строении пор является наличие торуса, способствующего активному перемещению воды из одной трахеиды в другую. В древесине или в коре многих хвойных имеются смоляные каналы или смоляные клетки, у некоторых видов смола начинает выделяться только как реакция на поранения. Листья с поверхности покрыты толстостенной эпидермой с мощной кутикулой; устьица глубоко погруженные. Под эпидермой располагается гиподерма, за ней толстый слой складчатого мезофилла со смоляными ходами, а в центре - пучок с камбием. По его периферии находится трансфузионная ткань, которая осуществляет связь между пучком и мезофиллом. Таким образом, у всех вечнозеленых хвойных хвоя имеет ксероморфную структуру.
Спороношение у хвойных наступает в разном возрасте; у светолюбивых видов - на 15-20-м году жизни, а у медленно растущих теневыносливых - на 40-60-80-м годах жизни. Мужские шишки, представляющие собой микростробилы, чрезвычайно разнообразны по характеру расположения, размерам и форме как самих шишек, так и микроспорофиллов (рис. 78). У наиболее древних родов (араукарии, агатиса, секвойи) мужские шишки одиночно располагаются на вершинах побегов или в пазухах листьев. Их размеры достигают 10-25 см и содержат сотни микроспорофиллов. Однако у большинства хвойных мужские шишки более мелкие - измеряются несколькими сантиметрами, у некоторых даже миллиметрами, а количество микроспорофиллов измеряется несколькими десятками. Как правило, шишки занимают пазушное положение, причем часто они образуют рыхлые или плотные скопления - констробилы. Прослеживающаяся эволюционная тенденция перехода одиночных крупных мужских шишек к многочисленным мелким имеет очень важное биологическое значение: во-первых, ускорение сроков образования пыльцы увеличивает вероятность опыления; во-вторых, автономность каждой мужской шишки повышает надежность их общей сохранности. Форма микроспорофиллов может быть весьма разнообразной - щитковидная, треугольная, ромбовидная, плоская. Редко они имеют радиальную симметрию, чаще они дорзивентральные. Одними авторами признается первичность радиальной структуры, другими - дорзивентральной, но вероятнее всего, те и другие структуры возникали независимо друг от друга, что подтверждается палеонтологическими данными. Дальнейшая их эволюция шла параллельными путями. Количество микроспорангиев, их размеры и характер прикрепления тоже весьма разнообразны. У более древних видов количество микроспорангиев может достигать 10-20 и они свободно свешиваются с нижней стороны спорофиллов. У более продвинутых видов количество их сокращается до 3-2; причем иногда (как, например, у сосны) они погружаются в ткань спорофиллов и срастаются с ней; этим достигается лучшая защита формирующейся пыльцы.
Женские шишки хвойных представляют собой констробилы. На их оси располагаются кроющие чешуи, в пазухах которых формируются семенные чешуи. На верхней (внутренней) стороне сидят семязачатки. Вопрос о происхождении семенной чешуи обсуждается учеными на протяжении десятков лет, причем разными учеными она интерпретировалась по-разному. На основании изучения палеоботанического материала, данных анатомии и сравнительной морфологии большинство современных ученых считают, что семенная чешуя - это сильно видоизмененный спороносный побег, т.е. мегастробил (рис. 79). В пользу этого имеется ряд аргументов. Во-первых, семенная чешуя расположена в пазухе кроющей чешуи, а по законам морфологии в пазухе листа может возникать только побег. Во-вторых, нередко у лиственницы и у некоторых других пород наблюдаются случаи прорастания женской шишки в удлиненный побег. При этом кроющие чешуи приобретают зеленый цвет и форму хвои, а в их пазухах развиваются укороченные вегетативные побеги.
В-третьих, среди самых древних, вымерших представителей семейства лебахиевых, описанных из верхнего карбона - перми, найдены репродуктивные органы. На их осях в пазухах кроющих чешуи располагались не семенные чешуи, а укороченные побеги. Каждый такой побег в основании нес несколько стерильных чешуи, а в верхней части имелось от 3 до 1 семязачатка на длинных ножках - семяножках. Однако у верхнепермских вольциевых в пазухах кроющих чешуи располагался пазушный комплекс промежуточного строения между побегом и семенной чешуей. У большинства современных хвойных женские шишки состоят из большого количества кроющих и семенных чешуи. У некоторых видов их может быть 3-4 (например, у можжевельника, у подокарпуса), а у видов подокарпуса и у тисса происходит сокращение до одной. Имеются многочисленные варианты в соотношении размеров кроющей и семенной чешуи, в их консистенции, форме. Для каждого семейства характерны свои особенности кроющих и семенных чешуи.
Мужской гаметофит - пылинка начинает формироваться внутри микроспорангия; он состоит из 1-3 быстро отмирающих проталлиальных клеток (у тисса и таксодиевых они не образуются), антеридиальной и сифоногенной клеток (см. рис. 66). У некоторых видов из спорангия вылетает одноклеточная непроросшая спора. Имеются интересные наблюдения, показывающие сложный путь, который проделывает пылинка внутри женской шишки, засасываясь и перемещаясь по аэродинамическим законам. Вихревые потоки, которые создаются внутри шишки, во многом определяются конфигурацией семенной чешуи, ее размерами. Путь, который проделает пылинка, также зависит от ее размеров, формы, удельного веса, поэтому пылинка другого вида практически не достигает семязачатков. Большую роль при этом играет наличие или отсутствие пыльцевых мешков, их количество. Улавливанию пыльцы семязачатками способствует выделяющаяся у многих видов сахаристая жидкость. Семязачатки хвойных, в отличие от саговниковых и гинкго, не формируют ни пыльцевых, ни архегониальных камер. Поэтому попадающая на семязачаток пылинка данного вида сразу начинает формировать пыльцевую трубку. У одних видов пыльцевая трубка в течение нескольких недель достигает архегония, а у других это прорастание длится несколько месяцев (например, у сосны), и оплодотворение происходит только спустя год после опыления. Пыльцевая трубка доставляет к архегонию 2 спермия, один из которых оплодотворяет яйцеклетку, а другой погибает. Обычно оплодотворяется яйцеклетка только одного архегония, поэтому в большинстве случаев в семени формируется лишь 1 зародыш, реже их 2. Зародыши хвойных у разных видов имеют от 2-3 до 15-17 семядолей. Семена всех хвойных прорастают после периода покоя; у одних видов оно подземное, у других - надземное.
В пределах подкласса хвойных выделяют 7 семейств, объединяемых одним порядком.
Хозяйственное значение сосен переоценить трудно. Древесина ее используется в жилищном строительстве, в кораблестроении, в вагоностроении, в авиационной промышленности, в столярном и мебельном производстве. Она высоко ценится у забойщиков для крепления в шахтах, так как при смещении пород она издает характерный скрип, предупреждающий об опасности. Смола, вытекающая из стволов сосны, - живица используется для изготовления канифоли, скипидара, камфоры. Хвоя, содержащая большое количество витамина С и каротина, идет на изготовление хвойно-витаминной муки, применяемой в животноводстве. Сосны, хотя и весьма декоративны, но оказываются очень чувствительными к загрязнению воздуха, поэтому для озеленения городов они не могут быть использованы.
Все голосеменные - растения разноспоровые, причем развитие мегаспоры в женский гаметофит - эндосперм происходит внутри мегаспорангия (нуцеллуса) на материнском организме. Следовательно, опыление и оплодотворение также осуществляются на материнском организме.
Появление интегумента привело в формированию семязачатков различных структур; причем для каждого таксона голосеменных характерен свой тип семязачатка.
В отличие от покрытосеменных у голосеменных отсутствует пестик, поэтому переносимая ветром пыльца непосредственно попадает на семязачаток.
Из семязачатка развивается семя, принципиально отличающееся от одноклеточной споры. Семя, являющееся зачатком нового организма, имеет зародыш, запас питательных веществ и надежную защиту в виде семенной кожуры. Это обстоятельство в значительной степени способствовало широкому распространению семенных растений и занятию ими господствующего положения на Земле.
Наличие зародышевого корешка семени послужило базой для формирования корневой системы стержневого типа, характерного для большинства древесных форм.
Наличие длительно функционирующего камбия, который у большинства голосеменных продуцирует мощную древесину, также явилось основой формирования древесных жизненных форм. Для голосеменных растений это направление оказалось генеральным.
Листья синтеломного происхождения эволюционировали у голосеменных в двух направлениях. У наиболее древних видов, имеющих неветвящиеся побеги, сохранились длительно живущие крупные листья с рассеченной листовой пластинкой. С появлением ветвления происходило уменьшение размеров листьев, что оказалось особенно важным в условиях холодного или сухого климата. В настоящее время голосеменные, представленные в основном хвойными породами, имеют довольно широкий ареал в Северном полушарии, и вместе с покрытосеменными на обширных территориях занимают ведущее положение в формировании растительных сообществ.
25-30 ОТДЕЛ ПОКРЫТОСЕМЕННЫЕ, ИЛИ ЦВЕТКОВЫЕ РАСТЕНИЯ (ANGIOSPERMAE, ANTHOPHYTA, ИЛИ MAGNOLIOPHYTA)
Наиболее крупный отдел растительного царства, насчитывающий более 350 семейств, 13 тыс. родов и до 240 тыс. видов. В настоящее время покрытосеменные распространены по всему земному шару - от тропиков до арктических и антарктических пустынь.
Автотрофные цветковые растения - важнейший активный компонент биосферы. От их жизнедеятельности зависит течение общебиосферных процессов обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования.
В наземных условиях цветковые обычно доминируют в растительном покрове, "каркас" большинства экосистем образуют именно они. Только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений - в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах - мохообразным, но и в таких случаях число видов покрытосеменных обычно превышает видовое разнообразие других отделов и подотделов растительного царства.
Понятно, что покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания животного населения. Животные связаны с ними самыми разнообразными трофическими, консортивными и иными связями. Многие группы животных вообще могли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Ясно, что и человек как биологический вид тоже не мог бы возникнуть и существовать, не будь на Земле покрытосеменных.
Покрытосеменные растения отличаются необычайным полиморфизмом, удивительной эволюционной пластичностью и могут существовать в самых разнообразных условиях обитания, вплоть до безводных пустынь, скал, солончаков, разбитых песков, шахтных терриконов, трещин в асфальте или бетоне. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду: в соленых водах морских мелководий вместе с водорослями растут десятки видов покрытосеменных.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания поражает воображение. Самый крохотный их представитель, ряска вольфия (Wolffla arrhiza) едва достигает 1- 1,5 мм в диаметре и похожа на свободно плавающую зеленую водоросль, а попросту - на булавочную головку. С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства Myrtaceae) вырастают высотой до 100 м, а длина побегов тропических лиан - ротанговых пальм измеряется сотнями метром. Преобладают среди покрытосеменных автотрофные растения, но немало среди них полупаразитов, паразитов и сапротрофов, полностью утративших хлорофилл и способность к фотосинтезу. Эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных - это общеизвестные примеры. А тончайшие приспособления к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов... Все это дает цветковым растениям возможность в полной мере реализовать свой эволюционно-адаптационный потенциал. Ни одна группа растений не могла выработать такого разнообразия приспособлений к определенным факторам среды.
В хозяйстве человека сложноцветные используются весьма многосторонне. Наиболее существенны следующие направления:
1. Декоративные растения. Самые разнообразные сложноцветные разводятся как красивоцветущие растения садов и цветников. Всемирную известность завоевали хризантемы и георгины. Многочисленные виды рода хризантема (Chrysanthemum) обитают в различных районах Старого Света. Уже задолго до н.э. они культивировались в Китае и скоро стали излюбленными цветами. Их роль в поэзии и живописи Китая огромна. Хризантема - национальный цветок Японии. Ее изображение можно видеть на гербах и печатях этой страны. Ко времени Великой французской революции хризантемы попали в Европу и скоро заняли господствующее положение в цветоводстве. Около 70 новых сортов выводят в Европе ежегодно. Особое значение имел так называемый далматский порошок, изготовляемый из корзинок хризантемы цинерариилистной (Ch. cinerariifolium). Он крайне эффективен против различных нежелательных домашних насекомых и, в противоположность синтетическим инсектицидам, определенно безвреден для теплокровных животных.
Род георгина (Dahlia), в диком состоянии обитающая в Мексике и Гватемале, также принадлежит к числу распространеннейших и излюбленных садовых растений. Выведено более 8000 сортов, главным образом махровых, у которых срединные трубчатые цветки превращены в ложноязычковые. Окраска, форма и размеры корзинок очень разнообразны, иногда превышают 20 см в диаметре. Большое значение имеет также астра садовая (Callistephus chinensis) и многие другие представители.
2. Лекарственные растения. Используются очень многие представители. Среди наших растений на первом месте, вероятно, стоит сушеница болотная (Gnaphalium uliginosum) - невзрачное растеньице с мелкими буроватыми корзинками, обитающее на полях, по берегам и днищам высыхающих луж и т.п. Применяют сушеницу прежде всего при язвенных болезнях желудка. Для полосканий, припарок, клизм часто используют ромашку лекарственную (Matricaria recutita). Из ноготков (Calendula officinalis) изготовляют популярную настойку для полосканий, а также мазь. Настойка арники, изготовляемая из корзинок и корневищ горного европейского вида арника горная (Arnica montana), - излюбленное гомеопатическое средство при различных поражениях. Особое применение имеет так называемое цитварное семя. Фактически это мелкие корзинки полыни цитварной (Artemisia cina), обитающей в полупустынях Средней Азии. Они исключительно богаты сапонином и применяются как глистогонное средство.
3. Овощные и масличные растения. Сюда относятся обыкновенный салат - латук (Lactuca sativa), у которого идут в кушанья молодые листья прикорневой розетки. Близкородственный вид - латук шиповатый (L. serriola) - обычный на хорошо освещенных мусорных местах, принадлежит к числу замечательных компасных растений. Листья у вида располагаются строго вертикально, причем плоскость пластинки ориентирована строго по меридиану. Таким образом, испарение сильно снижается. И в других родах у сложноцветных встречаются компасные растения.
В качестве наиболее известного заменителя кофе или добавления к нему в некоторых странах использовались измельченные корни цикория (Cichorium intybus), обычного на мусорных местах и особенно вдоль железных дорог крупного растения с голубыми корзинками. В послевоенные годы кофе с цикорием часто продавался в наших магазинах.
Исключительно важное значение имеет подсолнечник (Helianthus annuus) североамериканского происхождения. Растение давно попало в Европу и Россию, но лишь в XVIII в. стало известно, что семена его содержат много жиров и только в конце 1-й трети XIX в. научились отжимать масло. Произошло это в России, в Воронежской губернии. В России же возникли и первые сорта подсолнечника. Любопытно, что многие сорта, разводимые сейчас в Америке, русского происхождения. Помимо России, Украины и США особо большие посевы подсолнечника находятся в Румынии и Аргентине. Выведено множество сортов, высокорослых, используемых на силос, и низкорослых, удобных для машинной уборки. Диаметр корзинок иногда может достигать 60-65 см, а масличность у некоторых наших отечественных сортов - фантастической цифры 60%.
4. Каучуконосы. Многие сложноцветные содержат каучук в млечном соке или в паренхимных клетках. Довольно много каучука содержится в млечном соке - кок-сагыза (Taraxacum kok-saghyz) и тау-сагыза (Scorzonera tau-saghyz). В первые десятилетия советской власти, когда стремились ограничиться собственными ресурсами, им уделялось очень большое внимание. Еще большее значение имела гваюла (Parthenium argentatum) - невысокий кустарник родом из Мексики. Его выращивали и в промышленных масштабах, особенно во время второй мировой войны. Небольшие плантации гваюлы существовали и на юге нашей страны. В настоящее время каучуконосы - сложноцветные не имеют никакого значения. Качество их каучука, а тем более продуктивность, значительно ниже, чем у каучуковых деревьев из семейств тутовых и молочайных (см. выше, с. 281). Кроме того, возрастает роль синтетического каучука.
5. Сорные растения. Многие представители семейства - трудно искоренимые сорняки. В основном это многолетние травы, интенсивно размножающиеся корневыми отпрысками. В лесной зоне европейской части России наиболее важны осот розовый, или бодяк (Cirsium setosum), и осот желтый (Sonchus arvensis). У бодяка корневая система проникает на 4 м и даже глубже. Специальные корни, растущие сначала горизонтально, а затем загибающиеся вниз, развивают огромное количество побегов из придаточных почек, часто на большой глубине. Почки могут образоваться на изолированном участке корня всего 10 см длины. Бодяк, как и желтый осот, производит большое количество семянок, снабженных хохолками. В степной зоне большее значение приобретают другие виды - как латук, или молокан татарский (Lactuca tatarica), и горчак (Acroptilon picris). Последний - ядовитое растение. В живом виде он не поедается скотом, но в сене вызывает отравление.
Сложноцветные - одна из вершин в эволюции покрытосеменных. Такие их признаки, как соцветие - корзинка, имитирующее цветок, разнообразие цветков в одном соцветии в морфологическом и половом отношении, паппус, срастание пыльников и выметающие волоски, несомненно, можно рассматривать как высокопродвинутые.
Сложноцветные, по-видимому, родственны колокольчиковым. У последних нередко наблюдается аггрегация цветков в головчатые соцветия, несколько напоминающие корзинки сложноцветных. У некоторых срастаются пыльники и имеются выметающие волоски. Есть черты сходства и в биохимии. Не следует, конечно, забывать, что у колокольчиковых развивается нормальная чашечка, а гинецей их сильно отличается от гинецея сложноцветных.
Семейство магнолиевые (Magnoliaceae)
Деревья и кустарники с очередными цельными и цельнокрайними, реже лопастными листьями с прилистниками и крупными одиночными цветками. Цветки ациклические или спироциклические, обоеполые. Околоцветник простой, обычно венчиковидный, ярко окрашенный, из 6 или большего числа листочков, обычно в 3-членных кругах. Ось цветка сильно удлинена. Тычинок много, в неопределенном числе, часто мясистых, плоских с выраженным надсвязником. Стаминодии отсутствуют. Гинецей, как правило, апокарпный, из многих или нескольких плодолистиков, расположенных по спирали. Рыльца низбегающие. Семязачатки в различном числе вдоль брюшного шва, анатропные, с 2 интегументами. Плод из нескольких листовок или орешков, часто шишкообразный (рис. 111). Семена с эндоспермом, часто на длинных семяножках. Характерна мясистая ярко окрашенная саркотеста. Пыльца 1-бороздная. Узлы многолакунные.
10 родов и 220 видов, главным образом в горных субтропических лесах Юго-Восточной Азии, Меланезии, Северной и Центральной Америки.
Несмотря на небольшой объем (всего 220 видов!), магнолиевые относятся к числу наиболее известных семейств. Это связано не только с высокими эстетическими качествами магнолий и тюльпанных деревьев, но и с их ролью в филогенетических построениях.
Крупнейший в семействе род магнолия (Magnolia) распространен по всему ареалу семейства, заходя даже в районы умеренного климата. На южных Курильских островах (Кунашир) встречается как редкость магнолия обратнояйцевидная (М. obovata) - единственный дикорастущий вид семейства в России. В Крыму и на Кавказе часто разводят магнолию крупноцветковую (М. grandiflora), родина которой - юго-восток США. У этой магнолии крупные кожистые вечнозеленые листья и белые душистые, с исключительно сильным ароматом цветы до 20 см в диаметре. Листочков околоцветника 6, иногда 9 или 12. После цветения каждый из многочисленных пестиков превращается в листовку, тесно прирастающую брюшным швом к удлиненной оси цветка (см. рис. 111). Благодаря этому вскрывание листовок происходит по спинному шву. Очень эффектны ярко-красные ягодообразные семена, выставляющиеся на длинных семяножках.
Особого упоминания заслуживают прилистники. Они срастаются в некоторое подобие капюшона, выполняя важную защитную функцию не только для пазушных, но и для верхушечных почек.
Не все магнолии вечнозеленые растения, некоторые, как упоминавшаяся М. obovata, - листопадные деревья. Многие из них цветут перед распусканием листьев. Голые, без листьев, деревья, усыпанные крупными белыми, палевыми или ярко-розовыми цветами, необыкновенно красивы. Высокие декоративные качества магнолий давно прочувствованы японскими и китайскими художниками. Они заняли прочное место в восточно-азиатском искусстве.
Если у магнолий количество листочков околоцветника весьма различно и не всегда кратно трем, то у тюльпанного дерева (Liriodendron tulipifera) околоцветник строго 3-членный. Он состоит из 3 наружу отогнутых листочков и 6 вперед направленных и снабженных нектарниками, в 2 кругах. Впрочем, все листочки одинаково окрашенные, венчиковидные, а цветки схожи с тюльпанами (отсюда и название). Формулу такого околоцветника можно дать как K3 C3+3 или Kc3 CN3+3, отразив яркую окраску чашечки и наличие нектарников на лепестках. Как и у магнолий, в цветках тюльпанного дерева многочисленные тычинки и пестики сидят на удлиненной оси, однако плоды тюльпанного дерева своеобразны. Вместо листовок образуются орешки, причем верхняя стерильная часть каждого пестика преобразуется в крыло длиной несколько сантиметров. При созревании плода крыло все более и более оттопыривается от оси цветка, пока орешек не отрывается. При этом крыло действует как приспособление для полета и распространения семян.
Liriodendron tulipifera- мощное дерево высотой до 30 м, как и Magnolia grandiflora, родом с юго-востока США. Он разводится во многих местах Черноморского побережья Кавказа. Другой вид рода обитает в Юго-Восточной Азии. В отличие от магнолий листья у Liriodendron 4-лопастные. Из-за ряда своеобразных признаков (в дополнение к сказанному, отсутствие саркотесты) Liriodendron выделяют в особую трибу внутри семейства, противопоставляя всем остальным магнолиевым. Однако как и у всех магнолиевых, у него многолакунные узлы и 1-бороздная пыльца. Обычный на нашем Дальнем Востоке лимонник (Schisandra chinensis), ранее относившийся к магнолиевым, сейчас выделен из последних, прежде всего по 1-лакунным узлам и 3-6-бороздной пыльце.
Семейство магнолиевых часто рассматривается как очень примитивное, и сторонниками "раналиевой" доктрины и стробилярной гипотезы цветка ставится вместе со следующим семейством в основание системы цветковых растений. С этих позиций к несомненно примитивным признакам семейства принадлежат слабая дифференциация околоцветника и нестабилизированное число его членов, первичная полиандрия, листовидность и мясистость тычинок при выраженности надсвязников, апокарпный гинецей, низбегающие рыльца, удлиненная ось цветка, 1-бороздная пыльца, 3-лакунные узлы, сосуды с лестничной перфорацией, наличие волокнистых трахеид. Нельзя полностью исключить и филогенетические связи предков магнолиевых с беннеттитами (см. выше).
Семейство кувшинковые (Nymphaeaceae)
Водные укореняющиеся растения с плавающими или погруженными крупными округлыми черешковыми листьями и яркими крупными цветами. Цветки правильные, обоеполые, с простым или, чаще, двойным околоцветником. Чашелистиков 3-4 свободных или при основании несколько сросшихся. Лепестков обычно много, свободных в непостоянном числе. Встречаются переходные формы между лепестками и тычинками. Ось цветка удлинена, или плоская, или в виде гипантия. Тычинок много (до 700), часто листовидные, характерны надсвязники. Гинецей апокарпный, из многих плодолистиков, обычно погруженных в удлиненную ось цветка или прирастающих к бокаловидной оси (так называемая ложная ценокарпия). В последнем случае образуется общая рыльцевая поверхность из многих рылец - у рода кубышка (Nuphar). Завязь верхняя, полунижняя или нижняя; семязачатков много, висячих, анатропных, расположенных ламинально. Плоды - синкарпии, иногда ослизняющиеся или ягодообразные, без правильных способов вскрывания. Семена с арилусом. Эндосперм развит или отсутствует. Сосудистые пучки разбросаны (атактостелия). Сосуды отсутствуют. Характерны воздушные каналы, идиобласты, членистые млечники. 8 родов и 65-70 видов по всему свету.
Кувшинковые - очень характерное и широко распространенное семейство, хорошо приспособленное к водным местообитаниям. Пластинки их округлых или яйцевидных листьев и крупные цветки плавают на поверхности водоемов со стоячей или медленно текучей водой, однако растения могут укореняться на глубинах до 4 м! Наиболее обычны у нас кубышка (Nuphar luteum) с желтыми цветками и кувшинка, или водяная лилия (Nymphaea alba), с белыми цветками (рис. 117). Многие признаки у них общие; для обеих характерны постепенные переходы от тычинок к лепесткам. Но есть и различия. У кубышки околоцветник простой, венчиковидный, соответствующий по положению чашечке; вместо венчика у нее узкие нектарники, похожие на тычинки. А у кувшинки хорошо различимы чашечка и многолепестный венчик. Крупные белые цветки кувшинок необыкновенно красивы. Это и послужило причиной резкого сокращения численности растения. В настоящее время во многих местах принимаются меры по его охране.
Помимо N. alba к роду Nymphaea относится около 40 видов с белыми, розовыми, голубыми цветками. Все они очень декоративны. Сюда же принадлежит и священный цветок древних египтян - лотос египетский (N. lotus). Цветки его распускаются всегда вечером и закрываются на следующий день около 11 часов утра. Да и остальные виды рода цветут по часам, но в разное время. Так, цветы у нашей кувшинки открыты между 7 и 17 ч.
Во всех ботанических садах мира посетители стремятся увидеть тропические виктории (Victoria) родом и