 2013-12-31
2013-12-31 770
770Мониторинг природных популяций растений травянистых фитоценозов начал проводиться с 1986 года, т. е. сразу после катастрофы на ЧАЭС. В настоящее время анализируется более 2000 популяций растений, куда входит около 50 % травянистого состава Республики Беларусь. Популяции травянистых растений в 1986 году подверглись массовому острому облучению различными дозами, которое затем перешло в хроническое облучение, поэтому в последующий период и до настоящего времени растения постоянно подвергаются хроническому внешнему и внутреннему облучению. Более высокие дозы облучения растения получают при произрастании на почвах с высокой плотностью загрязнения радионуклидами.
Среди ответных реакций растений на воздействие ионизирующих излучений значительный интерес представляют нарушения морфогенеза. Фенотипические признаки того или иного вида растений являются показателем нормального прохождения многих тесно связанных между собой процессов онтогенетического развития. Разнообразные нарушения этих процессов приводят к большим или меньшим морфологическим отклонениям. Острое и хроническое облучение способно вызывать не только существенные отклонения в росте и развитии растений, но и способствует нарушению морфогенеза. Внешние эффекты, индуцируемые радиацией у растений, в большинстве случаев не отличаются от эффектов, вызываемых факторами нерадиационной природы. Поскольку ионизирующее излучение представляет собой сильный мутагенный фактор, то, вероятно, фенотипические отклонения могут быть генетически обусловленными. В то же время нарушение морфогенеза часто обусловлено поражением клеток меристемных тканей, которые характеризуются высокой радиочувствительностью.
При остром облучении растений в условиях эксперимента наряду с генетическими нарушениями выявлялись также морфологические и физиологические нарушения:
1) изменение формы органов (срастание или расщепление, ассиметрия или искривление, утолщение и др.);
2) стимуляция или угнетение процессов роста и развития;
3) изменение интенсивности опушения и цвета органов;
4) нарушение геотропизма;
5) нарушение порядка листорасположения и ветвления;
6) преждевременное опадение цветков и соцветий;
7) стерильность пыльцы, цветков, соцветий и растений;
8) опухоли органов;
9) гибель растений.
В результате мониторинга у травянистых растений естественных фитоценозов, произрастающих в условиях радиоактивного загрязнения после катастрофы на ЧАЭС, были также выявлены различные морфологические, физиологические и генетические отклонения. В 30-километровой зоне в популяциях травянистых растений наиболее часто наблюдались следующие морфологические отклонения, или радиоморфозы растений:
1) а номалии, обусловленные повреждением верхушечных и боковых почек;
2) снятие апикального доминирования и уменьшение высоты;
3) пробуждение спящих почек в узлах кущения и на побегах, способствующее усилению кущения и ветвления;
4) израстание и искривление главного и боковых побегов;
5) замедление или ускорение роста, т. е. карликовость или гигантизм. В популяциях растений наблюдалось поражение клеток верхушечных меристем стебля, что приводило к снятию апикального доминирования и изменению габитуса растений. Это выражалось в уменьшении высоты растений и длины междоузлий, в пробуждении спящих латеральных почек, в усилении кустистости и асимметрии ветвления побегов. Изменение габитуса растений вследствие подавления апикального доминирования в разные годы исследований наблюдалось у 80–90 % растений в популяциях полыни Маршала и золотой розги, у 30–60 % растений популяции ракитника русского и у 20–50 % растений популяций льнянки обыкновенной и ослинника двулетнего (приложение 5). Наиболее сильное проявление снятия апикального доминирования наблюдалось у видов рода подмаренник, где в результате поражения верхушечных меристем растения приобретали карликовость (их высота составляла всего 15–20 см при норме 80–100 см). Большинство таких карликовых растений не переходило в генеративную фазу.
У некоторых видов из латеральных почек формировались укороченные побеги, густо усаженные деформированными листьями и имеющие вследствие этого вид шишковидных образований. Эта аномалия была наиболее распространенной у полыни равнинной и составляла в разные годы 15–50 %. Израстание верхушек главного и боковых побегов наблюдалось у ракитника русского. Побеги представляли собой плотную розетку, сформированную ланцетовидными листоподобными образованиями, растения были ослаблены и погибали. Искривление побегов наиболее часто наблюдалось у букашника горного и вероники колосистой. Гигантизм растений был выявлен у ястребинки зонтичной и василька рейнского, а карликовость – практически у всех видов растений (5–10 % растений);
6) полимеризация – увеличение органов и их структур. Многие растения выделялись более крупными листьями, крупным выступающим жилкованием, а также увеличением толщины стеблей;
7) фасциация – срастание стеблей, соцветий и плодов. Фасциация стеблей выражалась в том, что они приобретали вместо округлой плоскую лентовидную форму (приложение 6). Эта аномалия встречалась нечасто. Она отмечалась в 1987–1990 годах у 20 % растений ракитника русского и у 2 % ястребинки зонтичной;
8) аномалии листьев – асимметрия, искривление, курчавость, махровость, морщинистость. Появление аномалий листьев объяснялось снижением интенсивности деления или гибелью меристемных клеток. Поэтому нарушалось или не происходило формирование определенных участков листовой пластинки. Аномалии листьев на загрязненной территории встречались практически у всех видов растений;
9) опухолевидное разрастание побегов. Опухолевидные образования на побегах наиболее часто встречались у ястребинки зонтичной (5 %), бодяка полевого (80 %);
10) аномалии при формировании генеративных органов и изменение формы и размера соцветий. У некоторых видов растений первые фазы онтогенеза проходили без видимых изменений, а повреждения проявлялись только при переходе растений в генеративную фазу. Наиболее ярко это проявлялось у вербейника обыкновенного, у которого верхняя плодоносящая часть побегов, вместо цветков и формирующихся из них клубочков, была густо усажена совершенно не характерными для этого вида узкими, деформированными, искривленными и асимметричными листьями, при этом края листьев были завернуты вниз. Аномалии генеративных органов у вербейника обыкновенного зависели от уровня загрязнения почвы и составляли от 5 до 50 %. Следует отметить, что вербейник относят к высокоплоидным видам (2n = 84), для которых характерна высокая пластичность и устойчивость к неблагоприятным воздействиям. Однако, к радиационному воздействию он оказался наиболее радиочувствительным среди других растений. Поэтому вербейник обыкновенный можно использовать в качестве биологического индикатора радиоактивного загрязнения. Аномальные израстания, возникающие на месте цветков, отмечались у дремы белой (60 %). У подмаренника настоящего наблюдались ветвистые, искривленные, а также совершенно бесформенные колосья. У подорожника настоящего за счет уменьшения длины центральных и боковых побегов часть стебля, несущая генеративные органы, приобретала более компактную и укороченную форму, при этом на стеблях формировались конусовидные или шаровидные соцветия. Аномалии при формировании генеративных органов этих растений даны в приложении 7;
11) спиралевидное закручивание корней. Спиралевидное закручивание корней было выявлено у бодяка полевого.
Из описанных радиоморфозов к наиболее распространенным относились кустистость, карликовость и гигантизм. Радиоморфозы наблюдались преимущественно у многолетних растений, что связано с накоплением повреждений в клетках и со слабым действием естественного отбора. У многолетних растений, как правило, большой набор хромосом в клетках и высокая способность к размножению.
Проведенный радиобиологический мониторинг природных популяций растений выявил, что фенотипические эффекты при хроническом облучении растений, произрастающих в условиях хронического облучения, имеют как различие, так и сходство с фенотипическими эффектами, обусловленными действием острого облучения. Морфологические аномалии растений являются следствием нарушения морфогенеза, в котором определенную роль играют строго упорядоченные в пространстве и времени процессы деления меристемных клеток. Под действием ионизирующих излучений постоянно делящиеся клетки меристем, которые характеризуются высокой радиочувствительностью, поражаются в разной степени (или погибают). При облучении происходит ослабление клеточных потоков, генерируемых меристемами, поэтому происходит снижение процессов обмена веществ и синтеза веществ, снижение ростовых процессов, что в конечном итоге приводит к изменению фенотипа растительного организма.
В популяциях травянистых растений также регистрировалось две волны гигантизма: первая волна – 1987–1988 годы, а вторая волна – 1991–1992 годы.
Кроме морфологических нарушений в популяциях естественных травянистых фитоценозов наблюдались следующие физиологические отклонения:
1) пигментация или хлороз побегов и верхушек цветоносов. Значительным проявлением хлороза побегов и верхушек цветоносов характеризовался бодяк (50 % растений), ракитник (30 %) и ежа сборная (10 %);
2) увядание растений и усыхание цветоносных побегов в жаркую погоду. Увядание растений наблюдалось у ястребинки (до 50 % растений) и у полыни Маршала (до 10 % растений), при этом увядающие верхушки цветоносных побегов полностью засыхали и не давали семян, что было связано с нарушением транспирации в жаркую погоду;
3) преждевременный опад листьев. Эта реакция растений на облучение была распространена у многих видов растений;
4) изменение окраски стеблей, листьев. Листья и стебли приобреталибледно-желтые и красноватые оттенки;
5) изменение активности ферментов, катализирующих процесс фотосинтеза. Под действием облучения у травянистых растений усиливается процесс поглощения углекислоты и фотосинтезирующая способность. В то же время может снижаться активность хлорофиллазы и усиливаться активность пероксидазы;
6) образование опухолей, вызванных бактерией Agrobakterium tumefaciens. Активное развитие таких опухолей обнаружено на загрязненных территориях у ястребинки зонтичной и ястребинки обыкновенной, а также у осота полевого, у которого опухолевидные ткани формировались у 80 % растений.
Динамика фенотипических аномалий в условиях хронического облучения сходна с динамикой генетических нарушений.
В 30-километровой зоне в популяциях травянистых растений выявлялись и генетические отклонения.
Мутантные эффекты растений сопровождались повышением частоты различных генных нарушений и хромосомных аберраций в половых клетках. При этом преобладала массовая фрагментация хромосом, что было обусловлено действием плотно ионизирующих излучений, т. е. облучением клеток и их структур альфа- и бета-излучением. В популяциях подорожника большого, подорожника ланцетолистного, кульбабы осенней, гипохереса укореняющегося, ястребинки зонтичной, тысячелистника обыкновенного, золотой розги, сивца лугового и других растений в результате трехлетнего мониторинга в 30-километровой зоне было выявлено отсутствие четко выраженной положительной зависимости частоты хромосомных нарушений и качества семенного материала от уровня радиационного фона. Например, при гамма-фоне до 10 мР/ч масса 1000 семян у подорожника большого практически не изменялась и составляла 178–194 мг. При гамма-фоне 15–20 мР/ч масса снижалась до 118 мг, а при 70–90 мР/ч – она возрастала до 250 мг. Аналогичная зависимость была установлена и для лабораторной всхожести семян. Увеличение массы 1000 семян и их всхожести при высоком радиационном фоне, по-видимому, связано с нарушениями в генеративной сфере, которые приводили к снижению завязываемости семян. При меньшем количестве завязавшихся семян происходит перераспределение пластических веществ, которые используются на формирование оставшихся неповрежденных или слабо поврежденных семян.
В то же время в 1986 году у некоторых популяций растений была выявлена положительная корреляция между величиной радиационного фона и уровнем мутаций. Например, у гипохереса укореняющегося при гамма-фоне 0,6; 1,4 и 5,3 мР/ч частота аберраций составляла соответственно 0,12; 1,64 и 2,98 %. В последующие годы такой связи не обнаруживалось, а выявлялась отрицательная корреляция.
Специфическая реакция на повышение гамма-фона была выявлена у кульбабы осенней, для которой характерен высокий уровень естественной мутационной изменчивости. В условиях повышенного радиационного фона у кульбабы осенней наблюдалось значительное повышение частоты мутаций (в 3–7 раз по сравнению с контролем), максимум которой приходился на 1988 год. К 1992 году произошло резкое снижение частоты мутаций до контрольного уровня. Для анализируемых популяций растений было характерно повышенное содержание клеток с множественными аберрациями хромосом. Клетки, в которых количество аберраций составляет более трех, были обнаружены у всех видов растений от 4 (подорожник ланцетолистный) до 17 % (сивец луговой), за исключением подорожника большого. В контроле такие клетки были выявлены только у двух видов – ястребинки зонтичной и сивца лугового, их количество составляло соответственно 5 и 10 %. Клетки, имеющие более пяти аберраций, также встречались в анализируемых популяциях, их количество составляло от 2 до 9 %. В контроле они были отмечены только у тысячелистника (0,7 %) и сивца лугового (3,0 %). Повышенное содержание клеток с множественными повреждениями хромосом обусловлено действием плотно ионизирующих излучений.
Анализируемые популяции растений характеризовались разной радиочувствительностью. Основываясь на теории «попадания и мишени» следовало ожидать, что повышенная частота хромосомных мутаций будет наблюдаться у видов растений с большим количеством хромосом и более крупными ядрами. Однако, при проведении исследований это не подтверждалось. Например, мицелис степной и ястребинка зонтичная имеют 2n = 18, а сивец луговой – 2n = 20 хромосом примерно одинаковой величины. А уровень индуцированных мутаций у этих растений составлял соответственно 1–5, 3–10 и 12–33 %. С другой стороны, гипохерес укореняющийся и мицелис степной по уровню мутаций различались незначительно, несмотря на то, что имели разный набор хромосом, составляющий соответственно 2n = 8 и 2n = 18, которые имели примерно одинаковые размеры. У тетраплоидной расы (2n = 36) и октаплоидной расы (2n = 72) тысячелистника обыкновенного частота хромосомных аберраций практически не изменялась и составляла соответственно 3–11 и 4–8 %.
Следует отметить, что динамика хромосомных аберраций и индуцированных мутаций не соответствовали динамике радиационного фона. В динамике радиационного фона выделяли два этапа:
1-й этап – 1986–1987 годы. Радиационный фон с высокого, который наблюдался в 1986 году, снизился к 1987 году в 1,5–3 раза, по причине распада короткоживущих радионуклидов;
2-й этап – 1987–1988 годы. Радиационный фон стабилизировался, и в последующем значительных тенденций к снижению не наблюдалось.
В динамике хромосомных мутаций выделяют три этапа:
1-й этап – 1986–1987 годы. Происходило нарастание частоты мутаций с максимальным проявлением в 1987 году;
2-й этап – 1988–1991 годы. Отмечен спад мутаций до уровня естественного мутирования;
3 этап – с 1992 года до настоящего времени. Наблюдается волновая кинетика мутаций, т. е. периодическое снижение или возрастание мутаций по сравнению с естественным мутагенезом.
Радиобиологические эффекты растений при хроническом облучении растянуты во времени и проявляются значительно позже, чем при остром облучении. Выделение в динамике хромосомных аберраций трех этапов объясняется следующими причинами. Внешнее облучение в 1986 году активизировало репарационную систему, поэтому радиопоражение было менее значительным. Дальнейший рост мутаций связан, во-первых, с возрастанием внутреннего облучения от инкорпорированных радионуклидов, во-вторых, с увеличением генетического «груза», в-третьих, с подавлением процессов репарации. Спад частоты мутаций во втором этапе вызван снижением радиационного фона и доз внешнего облучения. К этому времени в популяциях под действием естественного отбора произошло удаление радиочувствительных форм, которые формировали мутации. Волновая кинетика мутаций через 5–6 лет связана с возникновением и поведением радиочувствительных форм в популяции. При оптимальных условиях эти формы сохраняются, и за счет них повышается частота мутаций. При неблагоприятных условиях они удаляются естественным отбором, поэтому частота мутаций снижается до спонтанного уровня. Волновую кинетику мутаций характеризуют графики рис. 3.
Уровень хромосомных мутаций зависит от экологических особенностей вида или от ареала распространения. У видов с узким ареалом распространения частота спонтанных мутаций хромосом составляет примерно 0,5 %, а частота индуцированных мутаций, т. е. образовавшихся при облучении, в 6–8 раз выше и составляет 3–4 %. У видов с широким ареалом распространения частота спонтанных мутаций составляет 3–4 %, а частота индуцированных мутаций в 3–6 раз выше и составляет 9–24 %. При облучении популяций, представленных видами с широким ареалом распространения, хромосомные нарушения быстрее восстанавливаются до естественного уровня. Популяции многолетних растений более гетерогенны (или неоднородны) по многим признакам, в том числе и по радиочувствительности. У растений этих популяций более совершенна система репарации, а также более высокая адаптационная способность к изменяющимся неблагоприятным условиям среды обитания. В связи с этими особенностями популяции многолетних растений с широким ареалом распространения более радиоустойчивы.
Растения с узким ареалом распространения, а также растения, находящиеся на границе предела естественного распространения (редкие и исчезающие виды), характеризуются повышенной радиочувствительностью. На основании проведенного мониторинга было установлено, что в 30-километровой зоне из фитоценозов исчезли подорожник индийский, василек рейенский, полынь Маршала, сокращается численность ракитника и вербейника обыкновенного. Однако, популяции этих видов в фитоценозах занимали небольшое пространство и их исчезновение не нарушило нормального функционирования растительных экосистем.
В 1988 году в популяциях растений естественных фитоценозов не всегда наблюдалась прямая положительная зависимость между дозой и эффектом, что выявлялось при изучении комплекса показателей, характеризующих качество семенного материала (масса 1000 семян, энергия прорастания, всхожесть, начальные ростовые процессы).
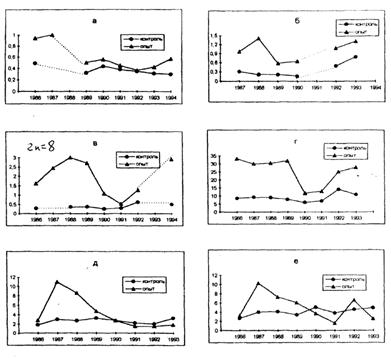
Рис. 3. Динамика хромосомных мутаций в природных популяциях растений
(по горизонтали – год исследования, по вертикали – клетки с аберрациями, %):
а – подорожник большой; б – подорожник ланцетолистный; в – гипохерес
укореняющийся; г – сивец луговой; д – кульбаба осенняя; e – ястребинка
зонтичная.
У большинства растительных объектов связь между мощностью дозы и качеством семенного материала отсутствовала или носила отрицательный характер. Отрицательная связь между дозой и массой 1000 семян, между дозой и лабораторной всхожестью была обнаружена у подорожника большого, синяка обыкновенного, клевера лугового и льнянки обыкновенной, а по ростовым реакциям (длина ростка и корешка) – у тысячелистника обыкновенного, петушиного проса и подорожника ланцетолистного. Причины отсутствия четко выраженной прямой зависимости между мощностью дозы и радиобиологическими эффектами в условиях хронического действия выбросов чернобыльской катастрофы разнообразны. Вероятно, основную роль играют, во-первых, радиационные факторы – действие комплексов радионуклидов, характеризующихся различной биологической эффективностью, внешнее и внутреннее облучение, во-вторых, экологические условия, комплекс которых специфичен для каждой природной популяции, способной модифицировать последствия радиационного воздействия, в-третьих, действие естественного отбора на разных стадиях эмбрионального и постэмбрионального развития, который удаляет радиочувствительные формы из популяции.
Через 15 лет после чернобыльской катастрофы среди природных популяций растений по радиочувствительности выделилось три группы растений. Радиочувствительность оценивалась по двум критериям: лабораторная всхожесть и частота хромосомных аберраций в клетках меристемы зародышевых корешков.
Первая группа – растения с повышенной чувствительностью к облучению. В эту группу входят многолетние растения – золотая розга, кульбаба осенняя, подорожник ланцетолистный, пазник укореняющийся, мицелис степной, а также двухлетнее растение – энотера.
Вторая группа – радиоустойчивые виды, у которых радиочувствительность не отличалается от контроля. В эту группу входят многолетние растения с широким ареалом распространения, у некоторых из них к этому времени уже произошла радиоадаптация – тысячелистник обыкновенный, полынь полевая, ромашка непахучая, дрема белая, тимофеевка луговая, василек луговой, вероника лекарственная.
Третья группа – растения с повышенной радиоустойчивостью. Сюда относится очень мало видов, например сивец луговой и полынь обыкновенная.
У некоторых видов растений был выявлен эффект радиоадаптации. Для выявления эффекта радиоадаптации проводилось дополнительное острое облучение семян более поздних послеаварийных репродукций. После этого производили разделение популяции на радиочувствительные и радиоустойчивые фракции. Радиоадаптация оценивалась по вышеуказанным критериям радиочувствительности.
В соматических и половых клетках растений облученных популяций, кроме структурных нарушений хромосом, были выявлены и количественные отклонения, среди которых чаще преобладала анеу- плоидия. Например, у ромашки непахучей в норме двойной набор хромосом составляет 36, т. е. 2n = 36. В облученной популяции у 18 % клеток количество хромосом изменялось и составляло 2n = 35, 2n = 37, 2n = 38.
Таким образом, природные популяции растений, подвергшиеся действию радиоактивных выбросов, характеризовались повышенным уровнем мутагенеза в первые послеаварийные годы и постепенным его снижением в последующие годы, отсутствием четко выраженной зависимости между мощностью дозы и вызываемым ею эффектом, а также различной радиочувствительностью видов.
В настоящее время доказано, что с ростом мощности поглощенной дозы в популяциях происходят следующие изменения: генные мутации, аберрации хромосом, видимые морфологические мутации, биохимические мутации, выпадение радиочувствительных видов, обеднение видового состава популяции. Нерегулярно наблюдаемые генетические эффекты регистрируются при мощности дозы 10-6–10-4 Гр/сут. Возрастание радиоустойчивости видов в популяции происходит при мощности дозы, составляющей 5 • 10-4 Гр/сут. При дальнейшем возрастании мощности поглощенной дозы происходит постепенное нарастание тяжести поражения и деградация популяции. Выпадение радиочувствительных видов наблюдается при мощности дозы 10-2 Гр/сут. Радиационные эффекты на экосистемном уровне начинаются с выпадения наиболее радиочувствительных видов, при этом в экосистеме происходит экологический сдвиг.
Таким образом, радиоактивное загрязнение природных экосистем, подвергающихся постоянному хроническому облучению, является новым экологическим фактором, вызывающим микроэволюционные изменения кариотипа в популяциях травянистых растений, а также морфологические и физиологические изменения растений.
В 1986 году морфологические, физиологические и цитогенетические изменения в популяциях агроценозов изучались в посевах озимой пшеницы, озимой ржи, ячменя, гороха и люпина в 30-километровой зоне и при плотности загрязнения почвы 37 кБк/м2 (1 Ки/км2). Посевы подверглись первоначально острому облучению в начальных фазах развития и хроническому облучению в течение всего периода вегетации.
В сортовых популяциях озимой пшеницы, озимой ржи и ячменя были выявлены следующие отклонения:
– замедление роста и развития растений;
– снятие апикального доминирования;
– повышенное кущение и образование боковых побегов;
– уменьшение размеров листьев, особенно флагового листа;
– изменение длины стебля (карликовость и гигантизм);
– хлорофилльные мутации и некрозы листьев;
– различные хромосомные аберрации с преобладанием фрагментов и мостов;
– увеличение продолжительности фаз онтогенеза и удлинение всего вегетационного периода;
– снижение активности фотосинтеза в 1,5–2 раза;
– стерильность пыльцы;
– плохая завязываемость семян.
Повышенная эффективность радиоактивных выбросов по сравнению с действием аналогичных доз внешнего, острого и хронического облучения от открытого источника цезия-137 выявлена на таких сельскохозяйственных культурах, как люпин, горох, озимая рожь, озимая пшеница и ячмень, при использовании тестов аберрации хромосом, хлорофилльных мутаций и ростовых реакций растений. В условиях радиоактивного загрязнения среды происходит нарастание уровня индуцированных мутаций во времени, несмотря на значительное снижение мощности экспозиционной дозы за счет распада короткоживущих радионуклидов. Увеличение частоты мутаций является следствием возрастания генетического «груза», обусловленного действием инкорпорированных радионуклидов, которое значительно выше действия внешнего облучения.
Одним из проявлений более высокой генетической эффективности радиоактивного загрязнения среды обитания являются множественные поражения хромосом, среди которых преобладает массовая фрагментация хромосом, которая была выявлена у пшеницы, ржи и ячменя. Установлено, что у ржи сорта Киевская 80 уровень хлорофилльных мутаций с 1986 по 1989 годы увеличился с 0,14 до 0,71 % (в контроле – 0,01 %), а у сорта Харьковская 60 – с 0,80 до 1,14 % (в контроле – 0,02 %).
Следует отметить, что данные о влиянии повышенного радиационного фона в результате последствий аварии на ЧАЭС не всегда однозначны и нередко противоречивы. Было выявлено отсутствие прямой зависимости между уровнем загрязнения и радиобиологическими эффектами, что не характерно при проведении экспериментов в контролируемых условиях. В ряде исследований четко прослеживалась линейная зависимость, в то время как в других только указывалось на наличие тенденции к положительной связи между мощностью дозы и эффектом. Иногда отмечались случаи обратной зависимости, т. е. относительно малые дозовые нагрузки вызывали больший эффект, чем более высокие. Например, у сортов ячменя Ролан и Жодинский при плотности загрязнения почвы 40 Ки/км2 было выявлено 9,1 и 11,8 % хромосомных аберраций в митозе. В то же время при плотности 7 Ки/км2 частота нарушений была выше и составляла 19,5 и 17,7 % соответственно. При этом в контроле частота хромосомных нарушений составляла 9,5 и 11,4 %. Аналогичные результаты были получены у ржи сорта Белта: контроль – 5,1 %, при плотности загрязнения почвы 40 Ки/км2 – 4,8 %, а при плотности 600 Ки/км2 – 2,4 %. У семян, выращенных при плотности загрязнения почвы 40 и 600 Ки/км2, снижалась лабораторная всхожесть и выживаемость растений, а у семян, выращенных при плотности загрязнения 7 Ки/км2, по этим показателям наблюдался стимуляционный эффект.
Посевы озимой ржи и озимой пшеницы, произрастающие на почве с плотностью загрязнения до 1000 Ки/км2 (37000 кБк/м2), характеризовались замедленным ростом и развитием, имели меньшую площадь всей поверхности листьев. Площадь флагового листа уменьшалась на 40–50 %. При меньшей плотности загрязнения радиоморфозов у этих зерновых культур было значительно меньше. У люпина при невысокой плотности загрязнения наблюдалось увеличение продуктивности растений и усиление процессов фотосинтеза, при этом происходила дезактивация основных ферментов, ответственных за фотосинтез: значительно снижалась активность фермента хлорофиллазы и усиливалась активность пероксидазы.
В посевах ячменя и пшеницы наряду с нормальными растениями было обнаружено много растений с радиоморфозами колосьев (приложение 8):
1) уменьшение и гигантизм главного колоса;
2) отсутствие остей на колосьях ячменя, а также повышенная неравномерная остистость, значительное увеличение остистости в верхней части колоса;
3) появление остей на колосьях пшеницы;
4) образование дополнительных колосков;
5) раздвоение и ветвление колоса;
6) стерильность и череззерница колосьев;
7) щуплость зерна и недоразвитость зародыша;
8) низкая всхожесть семян.
В посевах пшеницы, подвергшихся острому облучению в 1986 году, сформировались семена, из которых в последующие годы формировались растения, среди которых встречались мутантные формы колоса (отсутствие остей, выпадение отдельных колосков, раздвоение и ветвление колоса). Частота радиоморфозов в популяции озимой пшеницы на загрязненных радионуклидами территориях сохранялась на высоком уровне в течение нескольких поколений (табл. 7).
У нормальных семян этих культур технологические свойства зерна практически не изменялись. В деформированных зернах было более низкое содержание аминокислот, клейковины и белка. В семенах озимой пшеницы (Мироновская 808, Мироновская 27, Киянка, Полесская 70, Полесская 87, Безостая 1), озимой ржи (Саратовская и Харьковская 60) и инбредной линии кукурузы W64А, произраставших в 30-километровой зоне в 1987–1992 годах, накапливались фенольные соединения с измененным качественным составом.
Т а б л и ц а 7. Динамика радиоморфозов растений озимой пшеницы
| Тип морфоза | Год регистрации | ||
| Стерильность колосков в колосе | 49,0 | 29,8 | 1,9 |
| Укороченный колос | 10,0 | 9,4 | 0,8 |
| Расщепленный колос | 4,5 | 11,1 | 9,4 |
| Увеличение остистости колоса | 2,8 | 2,8 | 4,7 |
| Гигантизм колоса | 1,4 | 1,8 | 2,9 |
| Шероховатость остей | 1,4 | 3,4 | 2,9 |
| Дополнительные колоски | 14,0 | 14,8 | 29,7 |
| Изменение окраски стебля | 0,9 | 1,7 | 1,9 |
| Укороченный стебель | 4,5 | 5,7 | 4,9 |
| Удлиненный стебель | 4,4 | 4,7 | 5,4 |
На загрязненных территориях повышалось содержание антоцианов у лука-чернушки на 73 %, у фасоли овощной – 57 % и у кукурузы – 19 %. В чернобыльской зоне обнаружены ранее не описанные расы грибка стеблевой ржавчины (Puccinia craminis, Basidiomycetes). Устойчивость растений трех сортов пшеницы (Мироновская 808, Полесская 70 и Киянка), выращенных из семян, полученных в 30-километровой зоне, к стеблевой ржавчине в 2,6 раза ниже, чем в контроле.
На основании проведенного эксперимента было установлено, что облучение растений на загрязненных радионуклидами территориях оказывает более сильное воздействие на спорогенные клетки и пыльцу, чем экспериментальное гамма-облучение в контролируемых условиях, что отражено в табл. 8.
Т а б л и ц а 8. Частота аномальной пыльцы ячменя (на 1 млн. пыльцевых зерен) после 55 дней облучения на разном расстоянии от ЧАЭС и на экспериментальном гамма-поле (1992 год)
| Мощность дозы, мкЗв/ч | Доза, мЗв | Количество аномальных пыльцевых зерен, шт. |
| Облучение от чернобыльских радионуклидов | ||
| Облучение на гамма-поле | ||
| 3,0 | ||
| 29,6 | ||
При одинаковой мощности дозы, равной, согласно данным таблицы, 7, 515 мЗв/ч и 500 мЗв/ч, частота аномальных пыльцевых зерен при облучении от чернобыльских радионуклидов выше в 8,5 раза. Для выявления генетических эффектов семена, собранные в 1986 году с облученных растений, высевались в 30 километровой зоне и на чистой территории.
При этом было установлено, что до 70 % фенотипических изменений (радиоморфозов) передавалось по наследству до четвертого поколения. Массовая фрагментация хромосом выявлена не только у ряда представителей природной флоры (ослинника двулетнего, одуванчика лекарственного, подорожника ланцетовидного, кульбабы осенней, тысячелистника обыкновенного), но и у культурных растений агроценоза. В дальнейших поколениях, т. е. в четвертом – шестом поколениях, число нарушений варьировало, однако, в целом намечалась тенденция к нормализации у большинства линий.
Таким образом, изучение хронического действия выбросов Чернобыльской АЭС показало, что они способны вызывать существенные генетические изменения. При этом их мутагенные эффекты выше, чем эффекты эквивалентных доз хронического или острого облучения от точечных источников излучения. Наряду с повышенным выходом мутаций у растений из чернобыльской зоны выявлялись нарушения, которые обычно не отмечались при остром гамма-облучении в условиях экспериментов, что свидетельствует о более сильном поражении генетического материала. К таким нарушениям относят множественные аберрации хромосом, нарастание уровня мутаций во времени, несмотря на значительное снижение дозовых нагрузок и отсутствие адаптации у большинства популяций растений к действию радиации. Эти специфические явления обусловлены широким спектром излучений, испускаемых различными радионуклидами, и их совместным действием на растения. Увеличение генетических нарушений обусловлено также совместным действием внешнего и внутреннего облучения. Биологическая эффективность облучения за счет инкорпорированных радионуклидов значительно выше, чем от внешнего облучения в адекватных дозах, потому что радиобиологические эффекты растений зависят не только от дозы, но и от характера распределения дозы в микроструктурах органов и тканей, а также от способности радионуклидов включаться в критические структуры клетки.








