 2014-01-25
2014-01-25 990
990Рассматриваемая модель является модифицированным вариантом модели мышцы,впервые предложенной Хиллом. Данная модель включает в себя:СЭ –сократительный (контрактильный) элемент, ответственный за развитие силы тяги;.ПосУК-последовательный упругий компонент мышечных волокон. ПарУК – параллельный упругий компонент. ДЭ – демпфирующий элемент.
Последовательный и параллельный упругие компоненты представляют собой анатомически обособленные эластичные структуры, названные так в соответствии с их расположением относительно контрактильного элемента (СЭ.)
ПосУК- передает силу, развиваемую СЭ, на рычаг. Определяется упругостью актин- миозинового комплекса, обусловленного местами прикрепления актина к Z-дискам и местами соединения мостиков с активными центрами тонких нитей, а также сухожилиями Основная доля ПосУК сосредоточена в сухожилии. Так как жесткость сухожилия не зависит от активности контрактильного элемента, ее мож- но классифицировать как пассивную жесткость. Активная жесткость обусловлена количеством замкнутых мостиков, способных слегка растягиваться под нагрузкой.
Мышечная сила и активная часть ПосУК зависят от количества замкнутых мостиков. Следовательно,активная жесткость увеличивается с ростом напряжения мышцы...Если мышца пассивна, то ее напряжение поддерживается только за счет ПарУК. Зависимость удлинения ПосУК от развиваемой силы является экспоненциальной. Его максимальное удлинение, достигаемое при максимальном изометрическом напряжении мышцы, составляет от 2% до 7% от длины покоя мышцы..
ПарУК-параллельный упругий компонент. В него входят соединительно- тканные образования, составляющие оболочку мышечных волокон и пучков, а также механические свойства Т-системы и саркоплазматического ретикулуума..Следовательно, основную роль играют фасции и сарколеммы. Определенное влияние оказывают остаточные соединения актин- миозиновых мостиков. Механические свойства ПарУК проявляются при растягивании мышцы в расслабленном состоянии. Демпфирующеие свойства ПарУК не оказывают заметного влияния на силу тяги, а упругие начинают проявляться с равновесной длины мышцы, которая близка к длине покоя, при которой сократительный компонент развивает максимальную силу
Зависимость силы, развиваемой ПарУК. от его длины имеет экспоненциальный характер. В физиологическом диапазоне изменений суставных углов человека вклад указанной силы невелик.
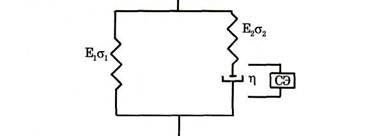
Рис. 2 Трехкомпонентная модель мышцы (СЭ – сократительный элемент)
СЭ – сократительный элемент представляет собой компонент, генерирующий силу. Его функционирование описывается кривыми «сила- скорость «и «сила- длина» В сократительный элемент входят актиновые и миозиновые филаменты, которые способны двигаться относительно друг друга за счет образования поперечных мостиков. Сила тяги СЭ определяется степенью перекрывания филаментов (величиной деформации мышцы.т.е. ее длиной), скоростью относительного движения нитей, композиционным составом мышцы, степенью возбуждения, температурой
Возбуждение мышцы определяется процессами активации каждого волокна и их рекрутированием...Зависимость силы, развиваемой ПарУК. от его длины имеет экспоненциальный характер. В физиологическом диапазоне изменений суставных углов человека вклад указанной силы невелик.
В мышцах рассеивается энергия; этот факт отражают в модели мышцы ведением ДЭ-демпфирующего элемента.
В заключение следует отметить, что упругость сухожилия примерно равна суммарной упругости всех мышечных волокон активной мышцы.
3 Структура поперечно – полосатой мышцы. Модель скользящих нитей.
Мышечная ткань представляет собой совокупность мышечных клеток (волокон), внеклеточного вещества (коллаген, эластин и др.) и густой сети нервных волокон и кровеносных сосудов.
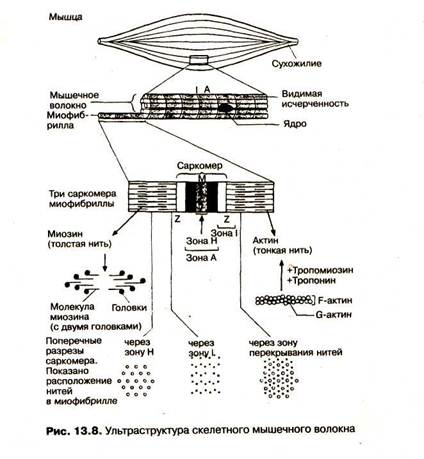
Поперечно-полосатая структура мышечных волокон может наблюдаться под обычным микроскопом. Отдельное мышечное волокно имеет диаметр 20-80 мкм и окружено плазматической мембраной толщиной 10 нм. Каждое отдельное волокно -это сильно вытянутая клетка. Длина отдельных волокон (клеток) может существенно варьироваться, в зависимости от вида мышцы, от сотен микрон до нескольких сантиметров. Внутри волокна, кроме известных органелл (ядро, ядрышко, митохондрии, аппарат Гольджи и др.), находятся сократительный аппарат клетки, состоящий из 1000 - 2000 параллельно расположенных миофибрилл диаметром 1-2 мкм, а также клеточные органеллы: саркоплазматический ретикулум и система поперечных трубочек - Т-система.
В миофибриллах различают (рис.): А-зону - темные полосы, которые в поляризованном свете дают двойное лучепреломление, то есть обладают свойством анизотропии (отсюда и название: А-зона), 1-зону - светлые полосы, не дающие двойного лучепреломления, то есть изотропные (отсюда название: 1-зона). В области I-зоны проходит темная узкая полоса — Z-диск (от нем. zwischenscheibe - промежуточный диск). Промежуток между двумя Z-дисками называется саркомером и является элементарной сократительной единицей мышечной клетки.
Саркомер - это упорядоченная система толстых и тонких нитей, расположенных гексагонально в поперечном сечении. Тол стая нить имеет толщину -12 нм и длину = 1,5 мкм и состоит \ из бёлка миозина. Тонкая нить имеет диаметр 8 нм, длину 1 мкм и состоит из белка актина, прикреплённого одним концом к Z-диску.
Актиновая нить состоит из двух закрученных один вокруг другого мономеров актина толщиной по 5 нм (рис. 3). Эта структура похожа на две нитки бус, скрученные^по_14_бусин в витке. В цепях актина регулярно примерно через 40 нм. встроены молекулы тропонина, а сама цепъ охватывает нить тропомиозина. При сокращении мышцы тонкие нити вдвигаются между толстыми. Происходит относительное скольжение нитей без изменения их длины. Этот процесс обусловлен взаимодействием особых выступов миозина - поперечных мостиков с активными' центрами, расположенными на актине. Мостики отходят от толстой нити пе риоди чно на расстоянии 14,5 нм друг от друга.
|
В расслабленном состоянии миофибрилл молекулы тропомиозина блокируют прикрепление поперечных мостиков к ак-тиновым цепям. Ионы Са активируют мостики и открывают участки прикрепления к актину В результате мостики миозина прикрепляются к актиновым нитям.,расщепляются молекулы АТФ и изменяется коформация мостиков: их головки поворачиваются внутрь саркомера. Это приводит к генерации силы,скольжению актина относительно толстой нити миозина к центру саркомера, что вызывает укорочение мышцы.Каждый цикл зымыкания –размыкания сопровождается расщеплением одной молекулы АТФ. Таким образом, актин-миозиновый комплекс является механохимическим преобразователем энергии АТФ.








