2018-02-13
2018-02-13 1007
1007Все клетки в той или иной мере чувствительны к механической деформации. Сокращение сердечной мышцы и гладких мышечных волокон усиливается при растяжении. Нервные волокна можно раздражать механическим путем, например дуновением. Деформация клеток меняет ионную проницаемость клеточных мембран. На этом явлении основана работа механорецепторов — клеток и систем, реагирующих на механическое воздействие— сжатие, растяжение, смещение. Простейшими механоре- цепторами являются свободные нервные окончания, лежащие либо между эпителиальными клетками, либо под ними. Кроме того, существует много специальных органов — луковиц, дисков, телец, спиралей, воспринимающих и передающих в нервные центры сигналы о механических воздействиях на ткань.
У рыб большое количество тактильных рецепторов находится в коже и плавниках. Механорецепторы находятся у рыб в сухожилиях, что позволяет рыбе регистрировать направление струй воды, касание предметов, анализировать свою позу. Характерных для высших позвоночных веретен, расположенных в мышцах, у рыб нет. При низком подпороговом воздействии электрический потенциал чувствительной клетки несколько повышается, но настоящие импульсные разряды появляются при превышении пороговой степени воздействия. Некоторые механорецепторы дают только один разряд в начале приложения раздражителя, другие дают пульсирующие разряды в течение всего периода механической деформации. Усиление воздействия, например, давления, растяжения учащает эти разряды. Упругие периодические колебания среды —звуки и вибрации — воспринимаются раз ными органами рыб (табл. 4).
Таблица 4. Упругие колебания и их восприятие рыбами

Инфразвуки или вибрации — колебания с частотой до 16 Гц — не воспринимаются рыбами во взвешенном состоянии без контакта с другими предметами и грунтом.
Механические раздражения, возникающие в результате гидродинамических процессов, воспринимаются органами боковой линии.
Звуковые колебания воспринимаются лабиринтовым аппаратом внутреннего уха.
Низкочастотные звуковые колебания воспринимаются рецепторами плавательного пузыря и боковой линии.
Основу органов боковой линии представляют специализированные волосковые клетки — нейромасты (рис. 21), располагающиеся на основной мембране. С помощью электронного микроскопа показано, что каждый нейромаст имеет различные виды волосков: один крупный — киноцилий и пучок из 40— 50 мелких — стереоцилий. Волоски погружены в желеобразную среду — купулу. Несколько нейромастов объединяются и образуют чувствительный бугорок. Кроме того, имеются опорные эпителиальные клетки. Наиболее просто органы боковой линии устроены у круглоротых и некоторых рыб (колюшка), у которых они представлены про- дольными рядами чувствительных клеток, расположенных на поверхности тела,— поверхностные органы. У акул и химер нейромасты расположены на дне желобка, проходящего по бокам тела, у ганоидов и большинства костистых рыб эти клетки располагаются на дне канала, сообщающегося с поверхностью тела небольшими тонкими ходами. Это канальные органы. У многих рыб органы боковой линии располагаются не только по бокам тела, но и в области головы, на жаберных крышках и нижней челюсти. Степень развития боковой линии у разных видов рыб различна. У верховки и корюшки она распространяется на небольшое расстояние позади головы, у других рыб она хорошо развита на всем протяжении туловища.
Раздражителями для чувствительных клеток боковой линии являются токи воды, вибрация и перепады давления, при этом происходит движение жидкости в каналах, приводящее к смещению волосков чувствительных клеток. Это воспринимается латеральными нервами туловища и головы. С помощью органов боковой линии рыба ориентируется в потоке воды, воспринимает движущиеся предметы, различает низкочастотные колебания.

Лабиринт, или внутреннее ухо, — парный орган, расположенный в хрящевой или костной капсуле височной части черепа. Лабиринт заполнен жидкостью — эндолимфой. У рыб он представлен тремя перепончатыми полукружными каналами и тремя отолитовыми органами — утрикулюсом, саккулюсом и лагеной (рис. 22). Строение и расположение полукружных каналов и других частей лабиринта у разных видов рыб различно.
Лабиринт делится на две части: верхнюю, к которой относятся полукружные каналы и утрикулюс, и нижнюю часть — саккулюс и лагена. Полукружные каналы расположены в трех взаимно перпендикулярных плоскостях. Каждый полукружный канал впадает в утрикулюс двумя концами, один из которых расширяется в ампулу. Здесь имеются возвышения, называемые слуховыми пятнами, или макулами, где расположены скопления чувствительных волосковых клеток. Тончайшие волоски этих клеток соединены студнеобразным веществом, образуя купулу. К волосковым клеткам подходят окончания VIII пары черепно-мозговых нервов. В утрикулюсе костистых рыб находится один крупный отолит. Отолиты располагаются также в лагене и саккулюсе

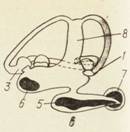
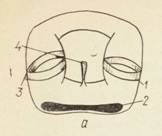
Рис. Внутреннее ухо или лабиринт рыб.
а — миксины; б — акулы; в — костистые рыбы; 1 — задняя криста; 2 — криста горизонтального канала; 3 — передняя криста; 4 — эндолимфатическнй проток; 5 — макула сак- кулюса; 6 — макула утрикулюса; 7 — макула лагены; 8 — общая ножка полукружных каналов
Отолит саккулюса используется для определения возраста рыб. Саккулюс хрящевых рыб сообщается с внешней средой посредством перепончатого выроста, у костистых рыб подобный вырост саккулюса заканчивается слепо.
У костистых рыб утрикулюс является основным рецептором положения тела. Отолиты с помощью студенистой массы соединены с волосками чувствительного эпителия. При положении головы теменем вверх отолиты давят на волоски, при положении теменем вниз висят на волосках, при боковых положениях головы отмечается разная степень натяжения волосков. С помощью отолитов рыба принимает правильное положение головы (теменем вверх), а следовательно, и туловища (спиной вверх). Для сохранения правильного положения тела также имеет значение информация, поступающая от зрительных анализаторов.

Рис. 24. Схематическое изображение веберова аппарата:
I-У — номера позвонков; 1 — перилимфатический проток; 2, 4, 6, 8 — связки и сроще- ння; 3 — стапес; 5 — инкус; 7—малеус; 9 — плавательный пузырь
Лабиринт связан с плавательным пузырем цепью веберовских косточек /каоПовые, обыкновенные сомы, харациновые, гнмнотовые), и рыбы способны воспринимать высокие звуковые тона (рис. 23, 24). С помощью плавательного пузыря происходит трансформация звуков высокой частоты в низкочастотные колебания (смещения), которые воспринимаются рецепторными клетками, у некоторых рыб, не имеющих плавательного пузыря, эту функцию выполняют воздушные полости, связанные с внутренним ухом.

Рис. 23. Расположение лабиринта, веберова аппарата и плавательного пузыря у карпа:
1— мозг; 2 — утрикулюс; 3 — саккулюс; 4 — объединительный канал; 5 — лагена: 6 —
перилимфатический проток; 7 — стапес; 8 — инкус; 9 — малеус; 10 — плавательный пузырь
Слуховой анализатор человека воспринимает колебания с частотой от 16 До 20 000 Гц. Звуки с частотой ниже 16 Гц относятся к инфразвукам, а выше 20000 Гц — к ультразвукам. Наилучшее восприятие звуковых колебаний наблюдается в интервале от 1000 до 4000 Гц. Спектр звуковых частот, воспринимаемых рыбами, по сравнению с человеком значительно сокращен. Так, например, карась воспринимает звуки в диапазоне 43—2760 Гц, карликовый сомик — 60—1600 Гц, акула S. canicula — 100— 2500 Гц. Рецепторы боковой линии воспринимают звуки на близком расстоянии, с помощью внутреннего уха происходит анализ звуков, доносящихся с большого расстояния.
рыбы могут определить и направление источника звука, но точно локализовать его не могут. Правильную локализацию источника звука они могут определить только на небольшом расстоянии с помощью боковой линии. Рыбы могут выделять нужные звуковые сигналы из шумов. Особенно хорошо они различичают однотонные звуки, которые подаются ритмично.
Центр, регулирующий работу акустико-латеральной системы, расположены в продолговатом мозге, он объединяет работу ядер VII, VIII и X пар нервов.