 2018-02-13
2018-02-13 1644
1644
Жизнь рыбы становится возможной благодаря наличию адекватных реакций ее организма на изменения, происходящие во внешней среде и в самом организме. Получение информации о физических и химических характеристиках среды называется рецепцией.
Структуры организма, воспринимающие эту информацию, называются рецепторами, т.е. приемниками.
- Рыба реагирует на световое излучение и внешний вид объектов материального мира, т.е. обладает зрением;
- реагирует на температуру окружающей среды, т.е. обладает терморецепцией;
- на ее поведение влияют многие растворенные в воде вещества, т.е. она обладает химической рецепцией — вкусом и обонянием;
воспринимает упругие колебания среды — слышит.
Рыба способна к проприорецепции — анализу своей позы и положения тела кроме того, она способна реагировать на ускорения, вызываемые смещением тела.
Легко обнаруживается тактильная рецепция рыб — реакция на прикосновения, удар, давление. На рыб оказывает влияние электрический ток, а также имеются указания, что в некоторых условиях она может реагировать на магнитное поле.
На радиоактивное излучение сублетальной интенсивности рыбы, как и другие живые существа, не реагируют.
ОРГАНЫ ЗРЕНИЯ
Главным органом зрения рыбы является глаз, хотя имеются и некоторые другие светочувствительные структуры, имеющие различную степень развития у разных видов. К восприятию света способна железа внутренней секреции — эпифиз.
Развитие зрения зависит от той роли, которую оно играет в жизни данного вида рыб. У пещерных рыб, миксин, личинок миног- глаза недоразвиты. У этих рыб хорошо развиты другие органы чувств — вкуса, осязания, орган боковой линии. У рыб, живущих в условиях хорошей освещенности (щука, окунь), глаза развиты хорошо, у придонных видов (вьюн, сом) они развиты хуже. Некоторые темнолюбивые виды (лещ, ерш, густера) имеют хорошо развитые глаза. Рыбы, живущие на больших глубинах, куда еще проникают лучи света, имеют огромные глаза улавливающие слабый свет, идущий сверху, и свечение, исходящее от живых организмов.
У глубоководных рыб могут быть большие глаза, но часто они недоразвиты.
В зависимости от расположения глаз различают рыб с монокулярным зрением — поля зрения двух глаз не перекрещиваются — и рыб с бинокулярным зрением — поля зрения двух глаз перекрещиваются. У большинства рыб глаза двигаются согласованно. У камбалы, морских игл и некоторых других рыб глаза могут двигаться независимо друг от друга.
Строение глаза рыб (СЛАЙД 7) в общих чертах напоминает строение глаза других позвоночных, в то же время глаза рыб имеют некоторые особенности, связанные с жизнью в водной среде. У рыб нет век, глаза их постоянно открыты. Лишь у некоторых акул имеется прозрачная мигательная перепонка, закрывающая глаз с внутреннего угла. У кефали и некоторых сельдей имеется жировое веко, частично закрывающее глаз. У рыб отсутствуют слезные железы, так как в отличие от наземных животных им нет необходимости смачивать глазное яблоко.
Строение глаза рыбы. Снаружи располагается склера, или белковая оболочка, которая впереди переходит в прозрачную роговицу. Склера состоит из плотной хрящевой ткани. Роговица представлена многослойным плоским эпителием, она прозрачна, только в краевой ее зоне имеются кровеносные сосуды. За склерой расположена серебристая оболочка, состоящая из нескольких слоев плоских клеток, содержащих кристаллы гуанина. Далее находится сосудистая оболочка, содержащая сосуды и пигментные клетки. В передней части глаза она переходит в радужную оболочку, в центре которой имеется отверстие — зрачок. Через зрачок световые лучи проникают внутрь глаза. Зрачок может расширяться и сужаться благодаря работе особых мышц, таким образом регулируется поступление в глаз света. Реакция зрачка на свет хорошо выражена у акул и угрей. К пигментному слою примыкает сетчатка. Отростки пигментных клеток могут укрывать светочувствительные клетки, так называемые палочки и колбочки. Палочки оценивают степень освещенности, силу света, колбочки осуществляют цветовосприятие. Палочки функционируют при дневном свете и в сумерках, колбочки — только при сильном дневном освещении.
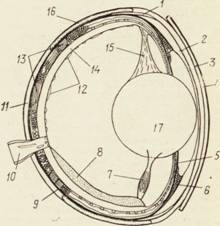
Рис. 6. Строение глаза рыбы: 1 — эпихороидальное лифматическое пространство; 2 — автохтонный слой; 3 — склероидный слой; 4 — дермальный слой (2,3 и 4 и составляют роговицу); 5 — серебристый слой радужины; 6— кольцевая связка; 7—мышца; 8 — серповидный отросток; 9 — серебристый слой сосудистой оболочки; 10 — зрительный нерв; 11 — белковая оболочка (склера); 12 — сосуды стекловидного тела; 13 хлоридная (сосудистая) оболочка; 14 сетчатка; 15 — подвеска; 16 — хрящевая склера; 17 — хрусталик
Пигментный слой уменьшает рассеяние света. За слоем светочувствительных клеток следует слой биполярных нервных клеток, а за ними идет слой ганглиозных клеток, аксоны последних дают начало зрительным нервам.
Там, где оптическая ось глаза пересекает сетчатку, расположено желтое пятно. Желтый цвет этой области зависит от присутствия каротиноидов. В центре желтого пятна расположено небольшое углубление — центральная ямка. На эту область сетчатки проецируются те объекты, которые находятся перед глазами и наиболее отчетливо видны. У дневных рыб здесь располагаются только колбочки. По периферии центральной ямки располагаются палочки, которые обусловливают периферическое зрение. Предметы, равно как цвета, проецирующиеся на эту область сетчатки, видны неотчетливо.
Палочки и колбочки состоят из внутреннего и наружного сегментов (члеников). Наружный сегмент содержит несколько сотен тончайших дисков он соединен с внутренним сегментом с помощью ножки. Внутренний сегмент содержит характерные для клетки органеллы.
Митохондрий образуют компактную массу — эллипсоид, который прилегает к наружному сегменту. Эллипсоид соединяется тонкими нитями протоплазмы (сократительный элемент — миоид) с ядром. Палочки имеют длинные цилиндрические наружные у колбочек наружные сегменты короткие, каплеобразные.
Наружные сегменты зрительных клеток являются их рецепторным аппаратом (рис. 7).
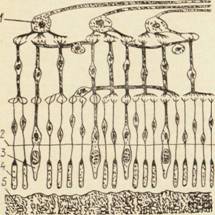
Рис. 7. Строение сетчатки глаза:
1— ганглиозные клетки; 2 — наружная пограничная мембрана, 3— колбочки; 4 — палочки; 5 — пигментный эпителий
У костистых рыб имеются колбочки трех видов: одиночные с коротким или длинным миоидом и близнецовые колбочки, для которых характерно тесное прилегание внутренних сегментов.
У рыб, амфибий и птиц в колбочках между внутренними и наружными сегментами расположена масляная капля, которая избирательно поглощает свет. Сетчатка позвоночных инвертирована (т.е. как бы; вывернута таким образом, что к. наружным сегментам фоторсцепторов свет доходит в последнюю' очередь). В эпифизе также имеется сетчатка с фоторецепторными: клетками, но она не инвертирована.
У светолюбивых рыб (окунь, форель, щука) светочувствительные элементы многочисленны с небольшим преобладанием палочек над колбочками. У этих рыб каждая фоторецепторная клетка контактирует с одной биполярной клеткой, а четыре биполярные клетки — с одной ганглиозной, что обеспечивает высокую остроту зрения при сравнительно низкой световой чувствительности. У рыб, избегающих яркого света (лещ. ерш, угорь, налим), в сетчатке в значительной степени преобладают палочки. У них с каждой биполярной клеткой связано до десяти и более фоторецепторных, а соответственно на одну ганглиозную клетку приходятся десятки и даже сотни фоторецепторных клеток. Это приводит к уменьшению остроты зрения, но к увеличению световой чувствительности. У некоторых видов морских глубоководных рыб колбочки вообще отсутствуют.
Доказано, что палочки пресноводных рыб содержат пурпурный зрительный пигмент—порфиропсин (максимум поглощения света приходится на, 520—540 нм); палочки морских рыб — родопсин (максимум поглощения света 500 нм) и смесь родопсина с порфиропсином (максимум поглощения света 500—540 нм); колбочки рыб содержат фиолетовый пигмент йодопсин (максимум поглощения света 662 нм) и голубой пигмент цианопеин (600 нм). Если рыбы из морской воды перемещаются в пресную, то родопсин у них меняется на порфиропсин.
Между палочками и колбочками имеются реципрокные отношения, т.е. когда функционируют палочки, колбочки заторможены, и наоборот.
У морских рыб светочувствительность сдвинута в коротковолновую часть. У пресноводных— в длинноволновую. В сумерки рыбы воспринимают коротковолновую часть спектра, днем — длинноволновую. Виды, живущиеют поверхностных слоях (керчак, морской налим), воспринимают более широкий спектр лучей света (400—750 нм), чем донные рыбы (скаты), воспринимающие более узкий спектр света (410—640 нм). В воде с увеличением глубины усиливается поглощение красных и ультрафиолетовых лучей, на глубине преобладают синие лучи, соответственно суживается воспринимаемый
спектр света.
Позади зрачка располагается хрусталик. Показатель преломления его у рыб равен 1,649—1,653. Показатели преломления роговицы и стекловидного тела приблизительно такие же, как у воды поэтому у рыб основную преломляющую функцию выполняет хрусталик.
Аккомодация — способность четкого видения предметов находящихся на разных расстояниях от глаза; у млекопитающих осуществляется за счет изменения кривизны хрусталика, у рыб имеется галлеров орган (специальные мышцы), за счёт которого изменяется положение хрусталика по отношению к сетчатке. Стекловидное тело расположено позади хрусталика и способствует сохранению формы глаза. При ранениях оно вытекает, глазные оболочки спадаются и глаз перестает видеть.
При попадании света на сетчатку в ней происходят фотохимические, химические, электрические и ретиномоторные процессы. К фотохимическим процессам относится распад родопсина, порфиропсина и йодопсина. Скорость распада родопсина и порфиропсина выше, чем йодопсина.
Колбочки возбуждаются при действии длинноволновой части спектра, а палочки — коротковолновой. Палочки примерно в 1000 раз более чувствительны к действию света, чем колбочки. При интенсивном освещении преобладают процессы распада зрительных пигментов, в темноте — восстановительные процессы. Иодопсин восстанавливается быстрее родопсина и порфиропсина. Распад и восстановление зрительных пигментов происходят в течение нескольких секунд. В сетчатке происходят электрические явления и процессы обмена веществ — гликолиз, распад и восстановление АТФ. Электрические явления заключаются в том, что при воздействии света в сетчатке возникают электрические потенциалы.
Ретиномоторные явления наблюдаются в сетчатке таких пойкилотермных животных, как рыбы и амфибии. На свету колбочки приближаются к наружной мембране, а палочки закрываются пигментными клетками, в темноте палочки подтягиваются к наружной мембране, а наружные сегменты колбочек прикрываются пигментными клетками. Движение фоторецепторных клеток происходит за счет расслабления и сокращения миоида, в пигментных клетках пигмент перемещается по отросткам (СЛАЙД 9).
Рецепторы сетчатки генерируют импульсы при существенном изменении их освещенности. При попадании на них света они дают импульс «на включение», при сильном уменьшении освещенности — «на выключение». Если освещенность не изменяется, то информация с сетчатки в мозг не поступает, и зрительные образы становятся невидимыми. Неподвижные предметы становятся видимыми потому, что движение глаз, как бы ощупывает изображение, особенно контуры предметов. Изображение на сетчатке перевернутое и уменьшенное. Движение воспринимается рыбами в результате так называемого киноскопического эффекта — последовательного перемещения зрительных образов с одних чувствительных элементов на другие.
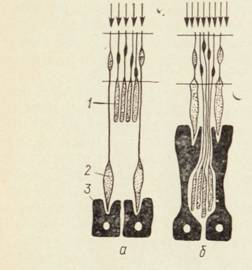
Рис.8. Схема ретинономоторной реакции.
а-слабое освещение, пигментный эпителий спался, светочувствительные элементыподтянуты ближе к свету; б-сильное освещение, светочувстветельные сегменты палочек упрятаны в пигментный слой; 1 – палочки; 2 – колбочки; 3 – пигментный эпителий.
В результате наличия рефрактерного периода (временной потери чувствительности после рабочего цикла) разрешающая способность ограничена. Критическая частота мельканий — вспышек света или движущихся в поле зрения полос — различается у разных видов рыб. Критическая частота мельканий у верховки 14 Гц, у атерины 67 Гц, а у человека 18—24 Гц.
Пространственное зрение (рис. 20), т.е. свойство восприятия предметов в пространстве, обеспечивается аккомодацией и остротой зрения. Последняя зависит от разрешающей способности преломляющих сред и сетчатки. Аккомодация глаза у рыб происходит за счет сокращений галлерова органа. При расслаблении галлерова органа глаз становится близоруким (миотропичным). Аккомодация лучше выражена у хищных рыб, чем у мирных. В спокойном состоянии глаза рыб гипермиотропичны, т.е. выражена установка «на бесконечность».
Острота зрения определяется наименьшим углом между двумя точками, при котором они воспринимаются раздельно.
Доказано, что многие костистые рыбы обладают цветным зрением. Скаты цвета не различают, т.е. являются ахроматами. У рыб, как и у человека, имеется три вида колбочек, воспринимающих три разных цвета. Одни колбочки реагируют на насыщенный красный цвет, другие на насыщенный зеленый, третьи — на насыщенный сине-фиолетовый цвет. В зависимости от количества и степени возбуждения колбочек воспринимаются различные оттенки цветов. Зрительные нервы содержат три группы афферентных нервных волокон, каждая из которых проводит раздражение от одного вида колбочек.
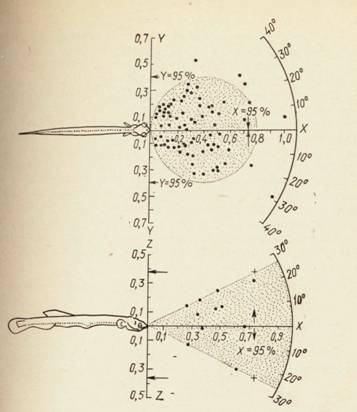
Рис. 20. Сектор охоты личинки анчоуса — пространственный масштаб (X, Y, 1 — в длинах тела личинки)








