2020-01-14
2020-01-14 202
202CRISPR (короткие палиндромные повторы, регулярно расположенные группами) — особые локусы бактерий и архей, состоящие из прямых повторяющихся последовательностей, которые разделены уникальными последовательностями (спейсерами). Спейсеры заимствуются из чужеродных генетических элементов, с которыми сталкивалась клетка (бактериофагов, плазмид). РНК, транскрибирующиеся с локусов CRISPR, совместно с ассоциированными белками Cas обеспечивают адаптивный иммунитет за счёт комплементарного связывания РНК с нуклеиновыми кислотами чужеродных элементов и последующего разрушения их белками Cas.
Локусы CRISPR могут выполнять функцию иммунитета только при наличии генов cas, которые обычно располагаются в непосредственной близости от CRISPR. Набор генов cas определяет тип системы CRISPR-Cas. Локусы CRISPR представлены короткими (обычно около 30—40 нуклеотидов длиной) прямыми повторами, которые отделяются друг от друга неповторяющимися спейсерами, произошедшими из ДНК тех чужеродных генетических элементов, с которыми сталкивалась клетка или её предшественники. Длина спейсеров обычно сопоставима с длиной повторов. Перед рядом повторов и спейсеров располагается лидерная последовательность, содержащая, как правило, промотор, с которого начинается однонаправленная транскрипция повторов и спейсеров CRISPR. Спейсеры полностью интегрированы в геном клетки и передаются её потомкам при делении. Стоит отметить, что у бактерий интеграция новых спейсеров в геном сочетается с утратой избыточных и чужеродных генов; поэтому бактериям удаётся избежать значительного увеличения размера генома. Кроме структурного сходства, различные системы CRISPR-Cas объединяют три ключевых этапа работы CRISPR-опосредованного иммунитета: приобретение, или адаптация, экспрессия и интерференция.
Системы CRISPR-Cas могут передаваться от микроорганизма к микроорганизму с помощью горизонтального переноса генов. Стоит отметить, что противодействие вторжению в бактерию чужеродных генетических элементов не всегда оказывается полезным для бактерии. Например, у бактерии Staphylococcus epidermidis может наблюдаться снижение устойчивости к антибиотикам, обусловленное уничтожением системой CRISPR-Cas тех конъюгативных плазмид, которые обеспечивали эту устойчивость. У Staphylococcus aureus пониженное количество локусов CRISPR приводит к увеличению числа профагов, плазмид и мобильных генетических элементов в клетке, что усиливает вирулентность бактерии.
Приобретение спейсеров. Поскольку опосредованный CRISPR приобретённый иммунитет закодирован в ДНК, процесс иммунизации включает копирование и вставку чужеродных генетических элементов в CRISPR в качестве новых спейсеров. Спейсеры составляют иммунологическую память, в которой хранится информация о прошлых инфекциях, и именно она лежит в основе ответа на повторное вторжение сходных генетических элементов. Большая часть данных о молекулярных механизмах приобретения новых спейсеров получена при изучении системы CRISPR I типа Escherichia coli и II типа Streptococcus thermophilus. Правильная ориентация и вставка нового спейсера происходит при участии последовательности, расположенной непосредственно выше первого повтора; таким образом, новые спейсеры добавляются к 5'-концу локуса CRISPR. Интеграция нового спейсера в промежуток между лидерной последовательностью и первым повтором осуществляется комплексом Cas1-Cas2-протоспейсер. У некоторых систем CRISPR-Cas в этом процессе участвуют дополнительные белки. При вставке нового спейсера происходит дупликация повтора, за счёт чего сохраняется правильная структура локуса, который должен начинаться с повтора. Поскольку спейсеры передаются от предков к потомкам при делении клеток, при наличии схожих спейсеров можно устанавливать филогенетические связи между штаммами, имеющими общие предковые спейсеры, а также штаммами, имеющими новые, недавно приобретённые спейсеры. У систем I и II типа может происходить вставка спейсера лишь от тех инородных элементов, у которых к протоспейсеру прилегает особая последовательность PAM (мотив, смежный с протоспейсером). Кроме того, бактерия должна отличать инородный генетический материал от своего, чтобы не вставить в качестве спейсера фрагмент собственной хромосомы и не нацелить систему CRISPR-Cas на свой геном, что было бы для клетки фатальным. Система CRISPR-Cas I типа E. coli отличает свою ДНК по наличию Chi-сайтов — 8-нуклеотидных мотивов, которые повторяются в её геноме в среднем каждые 5 тысяч пар оснований. Хотя из одного и того же инородного генетического элемента можно образовать множество спейсеров, в генетическом элементе некоторые мотивы оказываются при выборе будущего спейсера более предпочтительными. Вероятно, такие мотивы были зафиксированы в результате естественного отбора, связанного с эффективностью работы спейсеров; так, некоторые спейсеры дают начало crРНК, нацеливающим белки Cas и на частично комплементарные последовательности. Стоит отметить, что при столкновении с одним и тем же фагом разные клетки будут вставлять в качестве спейсера несколько отличающиеся фрагменты его генома, так что большие популяции, имеющие большое разнообразие спейсеров против одного и того же фага, оказывают более эффективное сопротивление: если фаг мутирует так, что один из имеющихся в популяции спейсеров станет неэффективен, то другие по-прежнему будут обеспечивать защиту.
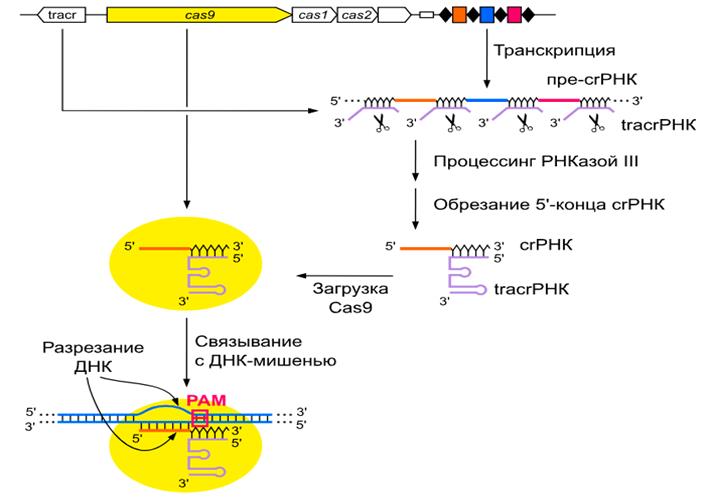
Экспрессия и образование crРНК После интеграции в CRISPR частей чужеродных генетических элементов требуется перевести их в форму, способную нацеливать белки Cas на последовательности-мишени для их распознавания и разрушения. Такой формой служит направляющая crРНК, которая содержит уникальную последовательность, комплементарную определённой мишени. Сначала ряд повторов и спейсеров CRISPR транскрибируется в единый длинный транскрипт — пре-crРНК, который далее разрезается на короткие crРНК. Большинство повторов в CRISPR являются палиндромами, поэтому соответствующие им участки пре-crРНК формируют шпильки. Во многих случаях именно эти шпильки распознаются белками Cas, процессирующими пре-crРНК в crРНК.
Как правило, транскрипция CRISPR зависит от лидерной последовательности и происходит постоянно, но с низкой скоростью. Однако скорость значительно увеличивается в стрессовых условиях или при столкновении клетки с фагами, обеспечивая ей быструю и эффективную защиту. Промоторные элементы были найдены не только в лидерной последовательности, но и в повторах. Несмотря на то, что за один раз может транскрибироваться весь локус, было показано, что некоторые спейсеры в локусе транскрибируются чаще других — в частности, таковы первые несколько спейсеров, располагающиеся после лидерной последовательности и первого повтора. Действительно, для клетки гораздо более выгодно иметь более сильную защиту от инвазивных элементов, с которыми она сталкивалась в недавнем прошлом, чем от тех, с которыми она встречалась давно.
Интерференция. На стадии интерференции crРНК связываются со своими мишенями за счёт спаривания оснований и, таким образом, направляют эндонуклеазы Cas на разрезание и разрушение мишени. Формирование комплекса crРНК и белков Cas обеспечивает эндонуклеолитическое разрушение комплементарных crРНК последовательностей НК. Хотя мишенями, в основном, являются двуцепочечные ДНК (дцДНК), некоторые системы CRISPR-Cas могут разрушать комплементарные одноцепочечные РНК (оцРНК). Системы CRISPR-Cas, распознающие дцДНК, требовательны по отношению к соседним с протоспейсером последовательностям: в частности, в системах типов I и II распознаются только мишени, содержащие мотив PAM (требование наличия PAM может служить для защиты от разрезания системой CRISPR-Cas клеточного генома). У систем, работающих с оцРНК, подобных требований нет. После начальной эндонуклеолитической атаки (внесения разрыва в мишень), производимой Cas, дальнейшее разрушение мишени может происходить под действием других нуклеаз