 2020-01-14
2020-01-14 5785
5785Строение глаза. Периферический отдел зрительного анализатора представлен глазом, который состоит из глазного яблока, расположенного в глазнице, и вспомогательных органов. Глазное яблоке имеет три оболочки (цвет. табл. VII, A),
Наружная оболочка состоит из двух частей: склеры и роговицы. Склера — плотная фиброзная оболочка, которую называют ещё белочной. Она непрозрачная и имеет характерный белый цвет, откуда происходит ее название. Роговица — передняя часть наружной оболочки, составляющая 1 / 6 всей ее поверхности. Она прозрачна и лишена сосудов. Роговица снабжена большим количеством чувствительных нервных окончаний, вследствие чего любое, самое незначительное раздражение ее поверхности вызывает защитные рефлексы слезотечения, моргания, смыкания век.
Среднюю оболочку глазного яблока называют сосудистой оболочкой.. Она состоит из трех частей: радужной оболочки, ресничного тела и собственно сосудистой оболочки. Радужная оболочка содержит пигмент, количество которого определяет цвет глаза. В центре ее расположено отверстие — зрачок. Величина его зависит от тонуса мышц радужной оболочки. Волоки мышцы, вызывающей сужение зрачка, окаймляют его по окружности, а мышцы, расширяющие зрачок, расположены радиально по отношению к нему. Мышца, расширяющая зрачок, иннервируется волокнами симпатической нервной системы, а суживающая — парасимпатической. Радужная оболочка, изменяя величину зрачка, регулирует тем самым количество поступающего в глаз света.
На уровне перехода роговицы в склеру радужная оболочка переходит в ресничное тело. На его внутренней поверхности, обращенной в полость глаза, находится около 70 ресничных отростков, которые венцом окружают хрусталик. Между ними и сумкой-капсулой, в которой расположен хрусталик, натянуты цинновы связки. В толще ресничного тела расположена мышца, при изменении напряжения которой меняется выпуклость хрусталика.
Ресничное тело переходит в собственно сосудистую оболочку, содержащую огромное число сосудов.
Внутренняя, очень сложная по строению оболочка глаза представлена сетчаткой. Она состоит из многих слоев клеток, различных по своему функциональному значению.
Позади радужной оболочки расположен хрусталик — прозрачное эластичное тело, имеющее двояковыпуклую форму. Пространство между хрусталиком и сетчаткой заполнено стекловидным телом, состоящим из прозрачного студенистого вещества. Между задней поверхностью роговицы и передней поверхностью радужной оболочки и хрусталика расположена передняя камера глаза, а пространство между задней поверхностью радужной оболочки и хрусталиком составляет заднюю камеру глаза. Обе камеры заполнены прозрачной жидкостью, содержащей некоторое количество белка и солей.
Вспомогательные образования глаза выполняют защитные и двигательные функции. Это веки, слезный аппарат и мышцы, осуществляющие поворот глазного яблока.
Проводниковый отдел зрительного анализатора начинается зрительным нервом, который направляется из глазницы в полость черепа. В полости черепа зрительные нервы образуют частичный перекрест, причем нервные волокна, идущие от наружных (височных) половин сетчаток, не перекрещиваются, оставаясь на своей стороне, а волокна, идущие от внутренних (носовых) половин ее, перекрещиваясь, переходят на другую сторону. После перекреста зрительные нервы называют зрительными трактами. Они направляются к среднему мозгу (к верхним холмикам четверохолмия) и промежуточному (боковое коленчатое тело и подушка зрительного бугра). Отростки клеток этих отделов мозга в составе центрального зрительного пути направляются в затылочную область коры головного мозга, где расположен центральный отдел зрительного анализатора. В связи с неполным перекрестом волокон к правому полушарию приходят импульсы от правых половин сетчаток обоих глаз, а к левому — от левых половин сетчаток.
Строение сетчатки. Самый наружный слой сетчатки образован пигментным эпителием. Пигмент этого слоя поглощает свет, вследствие чего зрительное восприятие становится более четким, уменьшается отражение и рассеивание света. К пигментному слою прилежат фоторецепторные клетки. Они состоят из ядросодержащей части, периферического и пресинаптического отростков. Периферические отростки из-за характерной формы называют палочками и колбочками. В соответствии с этим различают палочковые и колбочковые рецепторные клетки сетчатки. Они состоят из внешнего и внутреннего сегментов, соединенных так называемой ножкой.
Фоторецепторная клетка покрыта общей плазматической мембраной. Наружные сегменты палочковых и колбочковых клеток состоят из наложенных друг на друга дисков, число которых очень различно. Мембраны дисков представляют обычную липопротеиновую структуру, но отличаются от других клеточных мембран наличием зрительного пигмента, молекулы которого располагаются перпендикулярно к длинной оси палочковых и колбочковых клеток. Внутренний сегмент содержит большое количество митохондрий, плотно прилегающих друг к другу и образующих эллипсоид. В нем имеются различные ферменты, гликоген и РНК.
От ядросодержащей части палочковых и колбочковых клеток отходит центральный, пресинаптический отросток. Он заканчивается пуговчатым утолщением в палочковых и колбочковым в колбочках. Центральный отросток называют пресинаптическим в связи с; тем, что он образует синаптический контакт с дендритами биполярных клеток сетчатки.
Фоторецепторные элементы расположены на сетчатке неравно-! мерно. Глаз человека содержит 6—7 млн. колбочковых и ПО—125J млн. палочковых клеток.
На сетчатке имеется участок размером около 1,5 мм, который/ называют слепым пятном. Он совсем не содержит светочувствительных элементов. На 3—4 мм кнаружи от него находится желтое пятно, в центре которого расположено небольшое углубление — центральная ямка. В ней находятся только колбочковые клетки, а к периферии от нее число колбочковых клеток уменьшается и возрастает число палочковых. На периферии сетчатки имеются только
палочковые клетки.
За фоторецепторным слоем расположен слой биполярных клеток (рис. 59), а за ним слой ганглиозных клеток, который контактируют с биполярными. Oтростки ганглиозных клеток образуют зрительный нерв, содержащий около 1 млн. волокон. Один биполярный нейрон контактирую со многими фоторецепторами, одна ганглиозная клетка —» многими биполярными. Отсюда понятно, что импульсы от многих фоторецепторов сходятся одной ганглиозной клетке, ибо

Рис. 59. Схема соединения рецепторных элементов сетчатки с сенсорными нейронами:
1 — фоторецепторные клетки; 2 — биполярные клетки; 3 — ганглиозная клетка.
число палочек и колбочек превышает 130 млн. Лишь в области цен-: тральной ямки каждая рецепторная клетка соединена с одной биполярной, а каждая биполярная—с одной ганглиозной, что создает г наилучшие условия видения при попадании на нее световых лучей. - На сетчатке расположены еще горизонтальные и амакриновые клетки. Они связывают по горизонтали биполярные и ганглиозные клетки, что еще больше увеличивает число фоторецепторов, приходящихся на одну ганглиозную клетку.
Различия функции палочковых, колбочковых клеток и механизмы фоторецепции. Целый ряд факторов свидетельствует о том, чтопалочки являются аппаратом сумеречного зрения, т. е. функционируют в сумерки, а колбочки — аппаратом дневного зрения. Колбочки воспринимают лучи в условиях яркой освещенности. С их деятельностью связано восприятие цвета. О различиях в функциях палочковых и колбочковых клеток свидетельствует структура сетчатки различных животных. Так, сетчатка дневных животных — голубей, ящериц и др.— содержит преимущественно колбочковые клетки, а ночных (например, летучих мышей) — палочковые.
Наиболее отчетливо воспринимается цвет при действии лучей на область центральной ямки, если же они падают на периферию сетчатки, то возникает бесцветное изображение. При действии лучей света в наружном сегменте палочек зрительный пигмент родопсин разлагается на ретиналь — производное витамина А и белок опсин. На свету после отделения опсина происходит превращение ретиналя непосредственно в витамин А, - который из наружных сегментов перемещается в клетки пигментного слоя. Считают, что витамин А увеличивает проницаемость клеточных мембран.
В темноте происходит восстановление родопсина, для чего необходим витамин А. При его недостатке возникает нарушение видения в темноте, что называют куриной слепотой. В колбочковых клетках имеется светочувствительное вещество, сходное с родопсином, его называют йодопсином. Оно тоже состоит из ретиналя и белка опсина, но структура последнего неодинакова с белком родопсина.
Вследствие целого ряда химических реакций, которые протекают в фоторецепторных элементах, в отростках ганглиозных клеток сетчатки возникает распространяющееся возбуждение, направляющееся в зрительные центры головного мозга.
Цветовое зрение. Световые лучи с длиной волны от 0,4 до 0,8 мкм, вызывая возбуждение в колбочках сетчатки, обусловливают возникновение ощущения цветности предмета. Ощущение красного цвета возникает при действии лучей с наибольшей длиной волны, фиолетового — с наименьшей.
В сетчатке имеются три типа колбочек, реагирующих по-разному на красный, зеленый и фиолетовый цвет. Одни колбочки реагируют главным образом на красный цвет, другие — на зеленый, третьи — на фиолетовый. Эти три цвета были названы основными. Запись потенциалов действия от одиночных ганглиозных клеток сетчатки показала, что при освещении глаза лучами различной длины волны возбуждение в одних клетках — доминаторах — возникает при действии любого цвета, в других — модуляторах — только на определенную длину волны. При этом было выделено 7 различных модуляторов, реагирующих на длину волны от 0,4 до 0,6 мкм.
Оптическим смешением основных цветов можно получить все остальные цвета спектра и все оттенки.
Иногда наблюдаются нарушения цветовосприятия, в связи, с чем человек не различает тех или иных цветов. Такое отклонение отмечается у 8% мужчин и у 0,5% женщин. Человек может не различать один, два, а в более редких случаях все три основных цвета, так что вся окружающая среда воспринимается в серых тонах.
Адаптация. Чувствительность фоторецепторов сетчатки к действию световых раздражителей чрезвычайно высока. Одна палочка сетчатки может быть возбуждена при действии 1—2 квантов света. Чувствительность может меняться при изменении освещенности. В темноте она повышается, а на свету — уменьшается. Повышение чувствительности глаза в темноте называют темновои адаптацией, а понижение ее на свету — световой адаптацией. При темновои адаптации чувствительность может возрасти в 2 000 раз. Опыты показали, что адаптация зависит от влияний, приходящих из центральной нервной системы. Так, освещение одного глаза вызывает падение чувствительности к свету второго глаза, не подвергавшегося освещению. Предполагают, что импульсы, приходящие из центральной нервной системы, вызывают изменение числа функционирующих горизонтальных клеток. При увеличении их количества возрастает число фоторецепторов, соединенных с одно ганглиозной клеткой, т. е. возрастает рецептивное поле. Это и обеспечивает реакцию при меньшей интенсивности светового раздражения. При увеличении освещенности число возбужденных горизонтальных клеток уменьшается, что сопровождается падением чувствительности.
Построение изображения. Глаз имеет несколько преломляю1 сред: роговицу, жидкость передней и задней камер глаза, хруст лик и стекловидное тело. Построение изображения в такой системе очень сложно, ибо каждая преломляющая среда имеет свой ради кривизны и показатель преломления. Специальные расчеты показ ли, что можно пользоваться упрощенной моделью — редуцированным глазом и считать, что имеется только одна преломляющая поверхность — роговица и одна узловая точка (через нее луч пролетит без преломления), находящаяся на расстоянии 17 мм спереди от сетчатки (рис. 60).
Для построения изображения предмета АБ из каждой ограничивающей его точки берется два луча: один луч после преломлен проходит через фокус, а второй идет без преломления через уз вую точку (рис. 61). Место схождения этих лучей дает изображение точек А и Б — точки А1 и Б2 и соответственно предмет А1Б1. Изображение получается действительным, обратным и уменьшенным. Зная расстояние от предмета до глаза ОД, величин

Рис. 60. Расположение узловой точки Рис. 61. Построение изображения, и заднего фокуса глаза.
предмета АБ и расстояние от узловой точки до сетчатки (17 мм), можно вычислить величину изображения. Для этого из подобия треугольников АОБ и Л1Б1О1 выводится равенство отношений:

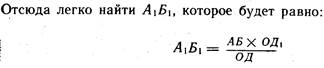
Преломляющую силу глаза выражают в диоптриях. Преломляющей силой в одну диоптрию обладает линза с фокусным расстоянием в 1 м. Для определения преломляющей силы линзы в диоптриях следует единицу разделить на фокусное расстояние в центрах. Фокус — это точка схождения после преломления параллельно падающих на линзу лучей. Фокусным расстоянием называют расстояние от центра линзы (для глаза от узловой точки) ho фокуса.
Глаз человека установлен на рассматривание дальних предметов: параллельные лучи, идущие от сильно удаленной светящейся точки, сходятся на сетчатке, и, следовательно, на ней находится фокус. Поэтому расстояние OF от сетчатки до узловой точки О является для глаза фокусным расстоянием. Если принять его равным 17 мм, то преломляющая сила глаза будет равна:

Аккомодация. Аккомодацией называют способность глаза видеть предметы на разном от него расстоянии. При аккомодации меняется кривизна хрусталика за счет сокращения ресничной мышцы (рис. 62). Она приближается к хрусталику, ослабевает натяжение цинновой связки, которая прикрепляется к его капсуле. Капсула расслабляется, и хрусталик, переставая испытывать ее давление, становится более выпуклым. Это сопровождается увеличением преломляющей силы и перемещением точки схождения лучей, идущих от ближнего предмета на сетчатку. Наименьшее расстояние ~ предмета до глаза, на котором этот предмет еще ясно видим,

Рис. 62. Механизмы аккомодации:
А — состойние покоя; Б — аккомодация: / — роговица; 2 — радужная оболочка; 3 — хрусталик; 4 — цииновы связки; 5 — ресничная мышца расслаблена; ё — ресничная мышца сокращена.
определяет положение ближней точки ясного видения, а наибольшее расстояние — дальней точки ясного видения. При расположении предмета в ближней точке аккомодация максимальна, в дальней — аккомодация отсутствует.
Разность преломляющих
сил глаза при максимальной аккомодации и при ее покое называют силой аккомодации. Величина аккомодации различна у разных людей. Она колеблется в зависимости от возраста. Максимальное ее значение составляет 14Д так что преломляющая сила глаза может возрасти до 73D: к 59D в покое прибавляется 14D, соответствующих силе аккомодации.
Рефракция. Преломляющие свойства нормального глаза называют рефракцией. Глаз без всяких нарушений рефракции, соединяющий параллельные лучи в фокусе на сетчатке, называют эммтропическим (рис. 63,Л). Если параллельно идущие лучи сходятся за сетчаткой, то глаз с таким отклонением от обычной рефракции называют гиперметропическим, или дальнозорким (рис. 63, Б). В этом случае плохо видимы близко расположенные предметы. Если же лучи сходятся перед сетчаткой, то глаз называют миопическим, близоруким (рис. 63, В). При таком нарушении рефракция плохо видимы далеко расположенные предметы.
Причина близорукости и дальнозоркости заключается в не-1 стандартной величине глазного яблока и в необычной преломляющей силе.
Большая величина переднезаднего диаметра глазного яблока, или большая преломляющая сила, сопровождается близорукостью малый диаметр, или малая преломляющая сила,— дальнозоркостью. При близорукости необходимы очки вогнутыми стеклами, которые рассеивают лучи; при дальнозоркости — с двояко выпуклыми.

Рис. 63. Схема рефракции:
А — нормальный глаз; Б — дальнозоркий глаз; В — близорукий глаз.
К отклонению рефракции относится также астигматизм. Он связан с тем, что преломляющие поверхности имеют разную кривизну в разных меридианах. Поэтому лучи, попадающие на них, неодинаково преломляются и сходятся не в одной точке. Этот недостаток в очень слабой степени присущ всякому глазу. Если посмотреть на рисунок 64, где одинаковые по толщине линии расположены горизонтально и вертикально, то одни из них кажутся более тонкими, другие — более толстыми. В тех случаях, когда эта аномалия рефракции сильно выражена, требуется подбор специальных цилиндрических стекол.
К особенностям преломляющих свойств любого глаза относится явление сферической аберрации.
Оно заключается в том, что лучи, проходящие через периферические участки хрусталика, преломляются сильнее, чем лучи, идущие через центральные его части (рис. 65). Поэтому центральные и периферические лучи сходятся не в одной точке. Однако эта особенность преломления не мешает ясному видению предмета, так как радужная оболочка не пропускает лучи и тем самым устраняются те из них, которые проходят через периферию хрусталика.
Неодинаковое преломление лучей разной длины волны называют хроматической аберрацией.
Острота зрения. Глаз человека воспринимает две точки пространства как раздельные даже при очень маленьком расстоянии между ними. Такую способность глаза называют остротой зрения. Острота зрения определяется наименьшим углом зрения, а следовательно, наименьшим расстоянием между рассматриваемыми точками, при котором они видны еще как раздельные. Для нормального глаза острота зрения определяется углом в 1'. Ему соответствует расстояние между изображениями точек на сетчатке в 5 мкм. Поскольку размер палочки колеблется от 2 до 5 мкм, принято

Рис. 64. Чертеж для выявления астигматизма.

Рис. 65. Схема, поясняющая явление сферической аберрации.
Лучи, падающие на периферию хрусталика и на его центральные участки, сходятся в разных точках А и Б.
считать, что для видения двух точек раздельными необходимо, чтобы между двумя возбужденными точками было не менее одной невозбужденной.
Практически для определения остроты зрения пользуются таблицами, в которых имеется несколько рядов цифр, букв или фигур, разных в каждом ряду по величине. Для получения относительной величины, характеризующей остроту зрения, следует расстояние, с которого испытуемый может без ошибок узнавать буквы или фигуры, разделить на расстояние, указанное возле данного ряда. Для нормального глаза острота зрения составляет единицу.
Бинокулярное зрение. При бинокулярном зрении предмет виден одиночным лишь в том случае, если его изображение возникает на идентичных участках обеих сетчаток. Идентичными точками сетчатки двух глаз называют области центральных ямок и все точки, расположенные от нее на одинаковом расстоянии и в одном и том же направлении (рис. 66). Остальные, несовпадающие точки сетчаток называют неидентичными. При попадании лучей на неидентичные точки изображение предмета оказывается раздвоенным.
Чтобы лучи от предмета попали на идентичные точки, необходимо сведение осей зрения на предмете. Сведение осей зрения на предмете называют конвергенцией. Конвергенция осуществляется путем вращения глазных яблок, которое происходит при сокращении шести наружных глазных мышц (рис. 67). Все мышцы, кроме нижней косой, прикрепляются к фиброзному кольцу, расположенному вокруг отверстия, из которого выходит зрительный нерв. Наружная прямая поворачивает глазное яблоко наружу, внутренняя — внутрь, верхняя прямая — вверх и одновременно наружу, нижняя прямая — вниз и внутрь. Верхняя косая мышца поворачивает глазное яблоко вниз и наружу, нижняя косая — вверх и наружу.

Рис. 66. Ход лучей при бинокулярном зрении:
А — фиксирование взором ближнего предмета; Б — фиксирование взором дальнего предмета; 1,4 -идентичные точки сетчатки; 2.3 — неидентичные (диспаратные) точки.
Рис. 67. Расположение глазных мышц:
I — верхняя прямая; 2 — верхняя косая; 3 — внутренняя прямая; 4 — нижняя прямая; 5 — нижняя
косая; 6 — наружная прямая.
Предметы, расположенные близко и далеко, нельзя одновременно отчетливо видеть. Если свести оси зрения на ближнем предмете, то дальний предмет будет при этом раздваиваться (рис. 66).
Зрение двумя глазами значительно облегчает восприятие пространства и глубины расположения предмета. Оценка расстояния до предмета может быть произведена и одним глазом. При рассматривании ближних предметов напрягается ресничная мышца; чем ближе предмет, тем сильнее ее напряжение. Степень напряжения мышцы дает представление о расстоянии до предмета. Способствует оценке расстояний также и то, что близкие предметы дают большее изображение на сетчатке, чем далекие. Однако наиболее точная оценка расстояния до тех или иных предметов пространства осуществляется при зрении двумя глазами. Чем ближе расположен предмет, тем сильнее должны быть повернуты глазные яблоки для сведения на нем осей зрения. Степень сокращения соответствующих мышц дает представление об удаленности предмета от глаза.
Зрение двумя глазами способствует определению формы предмета, его объема. При рассматривании предмета одна часть лучей, идущих от него, попадает на идентичные участки сетчаток, а другая — на неидентичные, но смещенные относительно друг друга на очень небольшое расстояние. Поэтому при рассматривании предмета поочередно то одним, то другим глазом мы видим его неодинаково: часть деталей видима и одним и другим глазом, часть только правым или только левым. Это дает представление об объемности предмета.
Строение слухового анализатора. Периферический конец слухового анализатора представлен ухом. Он делится на наружное, среднее и внутреннее ухо. Наружное ухо состоит из ушной раковины и наружного слухового прохода. Ушная раковина улавливает звуковые колебания, которые далее передаются по наружному слуховому проходу к барабанной перепонке. Наружный слуховой проход имеет длину около 24 мм. Он выстлан кожей с тонкими волосками и особыми потовыми железами, которые выделяют ушную серу.
Среднее ухо представлено барабанной полостью. В ней находится цепь слуховых косточек: молоточек, наковальня и стремя. Рукоятка молоточка срастается с барабанной перепонкой, а его головка образует сустав с наковальней, которая также соединяется суставом с головкой стремени.
На медиальной стенке барабанной полости находятся отверстия: окно преддверия (овальное) и окно улитки (круглое). Основание стремени закрывает окно преддверия, ведущее в полость внутреннего уха, а окно улитки затянуто вторичной барабанной перепонкой.
Барабанная полость соединяется с носоглоткой посредством слуховой, или евстахиевой, трубы. Через нее из носоглотки в полость среднего уха попадает воздух, благодаря чему выравнивается давление на барабанную перепонку со стороны наружного слухового прохода и барабанной полости. Когда затруднено прохождение воздуха по слуховой трубе,(воспалительный процесс), то преобладает давление со стороны наружного слухового прохода, и барабанная перепонка вдавливается в полость среднего уха. Это приводит к значительной потере возможностей барабанной перепонки совершать колебательные движения в соответствии с частотой звуковых волн.
Внутреннее ухо называют еще лабиринтом. Оно расположено в пирамиде височной кости. Различают костный и перепончатый лабиринты. Стенки костного лабиринта образованы костной тканью височной кости. Внутри костного лабиринта, в основном повторяя его форму, расположен перепончатый лабиринт. Между костным и перепончатым лабиринтами имеется перилимфатическое пространство, в котором находится жидкость — перилимфа.
Лабиринт делится на вестибулярный аппарат (орган равновесия) и улитку, которая относится к слуховому анализатору. Улитка — это костный канал, который образует 2,5 завитка вокруг стержня. Стержень улитки состоит из губчатой костной ткани, между балками которой расположены нервные клетки, образующие спиральный ганглий (рис. 68, А). От стержня отходит в виде спирали тонкий костный листок, состоящий из двух пластин, между которыми проходят миелинизированные дендриты нейронов спирального ганглия. Верхняя пластина костного листка
переходит в спиральную губу, или лимб, нижняя — в спиральную основную, или базиллярную, мембрану, которая простирается до наружной стенки улиткового канала. Плотная и упругая спиральная мембрана представляет собой соединительнотканную пластинку, которая состоит из основного вещества и коллагеновых волокон — струн, натянутых между спиральной костной пластинкой и наружной стенкой улиткового канала. У основания улитки волокна более короткие. Их длина составляет 104 мкм. По направлению к вершине длина волокон увеличивается до 504 мкм. Общее их число составляет около 24 тыс. Ширина всей базиллярной мембраны составляет у вершины 0,5 мм, а у основания, вблизи овального окна,— 0,04 мм.
От костной спиральной пластинки к наружной стенке костного канала под углом к спиральной мембране отходит еще одна мембрана, менее плотная — вестибулярная, или рейснерова.
Полость канала улитки разделена мембранами на три отдела: верхний канал улитки, или вестибулярная лестница, начинается от

Рис. 68. Схема строения внутреннего уха:
А — поперечный разрез улитки; Б — кортиев орган:1 — вестибулярная лестница; 2 — рейснерова мембрана; 3 — улитковый проток; 4 — сосудистая полоска; 5 — кортиев орган; в — барабанная лестница;
[т — спиральный ганглий; 8 — лимб; 9 — текториальная мембрана; 10— наружные волосковые клетки; III — внутренние волосковые клетки; 12 — основная мембрана; 13 — дендриты нейронов спирального ганглия.
окна преддверия; средний канал улитки — между вестибулярной и спиральной мембранами и нижний канал, или барабанная лестница, начинающаяся от окна улитки. У вершины улитки вестибулярная и барабанная лестницы сообщаются посредством маленького отверстия — геликотремы. Верхний и нижний каналы заполнены перилимфой. Средний канал — это улитковый проток, который тоже представляет собой спирально извитый канал в 2,5 оборота. На наружной стенке улиткового протока расположена сосудистая полоска, эпителиальные клетки которой обладают секреторной функцией, продуцируя эндолимфу. Вестибулярная и барабанная лестницы заполнены перилимфой, а средний канал — эндолимфой.
В улитковом протоке на основной мембране расположены чувствительные волосковые клетки, образующие звуковоспринимающий спиральный орган, или кортиев (рис. 68, Б).
Различают внутренние и наружные волосковые клетки. Внутренние волосковые клетки несут на своей поверхности от 30 до 60 коротких волосков, расположенных в 3—5 рядов. Число внутренних волосковых клеток составляет у человека около 3500. Наружные волосковые клетки расположены в три ряда, каждая из них имеет около 100 волосков. Общее число наружных волосковых клеток составляет у человека 12—20 тыс. Наружные волосковые клетки более чувствительны к действию звуковых раздражителей, чем внутренние. Волокна клеток спирального ганглия образуют синаптические контакты с внутренними и наружными волосковыми клетками. Для волосковых клеток установлено наличие и эфферентной иннервации, что обеспечивает координацию в работе правого и левого уха.
Над волосковыми клетками расположена текториальная мембрана. Она имеет лентовидную форму и желеобразную консистенцию. Ее ширина и толщина увеличиваются от основания улитки к вершине. Информация от волосковых клеток передается по дендритам клеток, образующих спиральный узел. Второй отросток этих клеток — аксон — в составе преддверно-улиткового нерва направляется к стволу мозга и к промежуточному мозгу, где происходит! переключение на следующие нейроны, отростки которых идут в] височный отдел коры головного мозга.
Механизм передачи и восприятия звука. Звуковые колебания улавливаются ушной раковиной и по наружному слуховому проходу передаются барабанной перепонке, которая начинает колебаться соответствии с частотой звуковых волн. Колебания барабанной перепонки передаются цепи косточек среднего уха и при их участив мембране овального окна. Колебания мембраны окна преддверия передаются перилимфе и эндолимфе, что вызывает колебания основной мембраны вместе с расположенным на ней кортиевым opraном. При этом волосковые клетки своими волосками касаютс текториальной мембраны, и вследствие механического раздражен* в них возникает возбуждение, которое передается далее на волокна; преддверно-улиткового нерва.
Слуховой анализатор человека воспринимает звуковые волны частотой их колебаний от 20 до 20 тыс. в секунду. Высота тона определяется частотой колебаний: чем она больше, тем выше по тону воспринимаемый звук. Анализ звуков по частоте осуществляется периферическим отделом слухового анализатора. Под влиянием звуковых колебаний прогибается мембрана окна преддверия, смещая при этом какой-то объем перилимфы. При малой частоте колебаний частицы перилимфы перемещаются по вестибулярной лестнице вдоль спиральной мембраны по направлению к геликотреме и через нее по барабанной лестнице к мембране круглого окна, которая прогибается на такую же величину, что и мембрана овального окна. Если же действует большая частота колебаний, возникает быстрое смещение мембраны овального окна и повышение давления в вестибулярной лестнице. От этого прогибается спиральная мембрана в сторону барабанной лестницы и реагирует участок мембраны, обладающий наименьшей жесткостью. Жесткость спиральной мембраны повышается от основания улитки к ее вершине. Поэтому при действии очень большой частоты прежде всего реагирует участок мембраны вблизи окна преддверия. При повышении давления в барабанной лестнице изгибается мембрана круглого окна, основная мембрана благодаря своей упругости возвращается в исходное положение. В это время частицы перилимфы смещают следующий, более инерционный участок мембраны, и волна пробегает по всей мембране. Колебания окна преддверия вызывают бегущую волну, амплитуда которой возрастает, и максимум ее соответствует какому-то определенному участку мембраны (рис. 69). По достижении максимума амплитуды волна затухает. Чем выше высота звуковых колебаний, тем ближе к окну преддверия находится максимум амплитуды колебаний спиральной мембраны. Чем меньше частота, тем ближе к геликотреме отмечаются наибольшие ее колебания.

Рис. 69. Место возникновения максимальных колебаний основной мембраны:
А — при действии высоких звуков; £ — при действии средних звуков; В — при действии низких звуков; 1 — овальное окно; 2 — круглое окно; 3 — бегущая волна; 4 — основная мембрана; 5 — геликотрема.
Установлено, что при действии звуковых волн с частотой колебаний до 1000 в секунду в колебание приходит весь столб перилимфы вестибулярной лестницы и вся спиральная мембрана. При этом их колебания происходят в точном соответствии с частотой колебаний звуковых волн. Соответственно в слуховом нерве возникают потенциалы действия с такой же частотой. При частоте звуковых колебаний свыше 1000 колеблется не вся основная мембрана, а какой-то ее участок, начиная от окна преддверия. Чем выше частота колебаний, тем меньший по длине участок мембраны, начиная от окна преддверия, приходит в колебание и тем меньшее число волосковых клеток приходит в состояние возбуждения. В слуховом нерве в этом случае регистрируются потенциалы действия, частота которых меньше частоты звуковых волн, действующих на ухо, причем при высокочастотных звуковых колебаниях импульсы возникают в меньшем числе волокон, чем при низкочастотных колебаниях, что связано с возбуждением лишь части волосковых клеток. Значит, при действии звуковых колебаний происходит пространственное кодирование звука. Ощущение той или иной высоты звука зависит от длины колеблющегося участка основной мембраны, а следовательно, от числа расположенных на ней волосковых клеток и от места их расположения. Чем меньше колеблющихся клеток и чем ближе они расположены к окну преддверия, тем более высоким воспринимается звук.
Колеблющиеся волосковые клетки вызывают возбуждение в строго определенных волокнах слухового нерва, а значит, и в определенных нервных клетках головного мозга.
Сила звука определяется амплитудой звуковой волны. Ощущение интенсивности звука связано с различным соотношением числа возбужденных внутренних и внешних волосковых клеток. Поскольку внутренние клетки менее возбудимы, чем внешние, возбуждение большого числа их возникает при действии сильных звуков.
Электрические явления в улитке. В улитке без всякого звукового раздражения можно зарегистрировать мембранный потенциал волосковых клеток и потенциал эндолимфы. Мембранный потенциал волосковых клеток составляет —70 мВ, а потенциал эндолимфы -80 мВ. Отсюда следует, что на границе волосковых клеток с эндолимфой имеется большой потенциал, равный 80—(—70) = 150 Под влиянием положительного. потенциала эндолимфы мембрана волосковых клеток становится очень нестабильной и при их соприкосновении с текториальной мембраной дает резкие изме-1 нения МП. Сдвиг в величине МП вызывает возникновение электрического эффекта, который называют микрофонным потенциалом. Он проявляется в электрических колебаниях, которые по частоте соответствуют звуковым колебаниям и которые можно зарегистрировать от любой из лестниц улитки. Микрофонный потенциал связан с возбуждением волосковых клеток и исчезает при их разрушении. Наибольшая величина его отмечается вблизи кортиева органа, поэтому считают что микрофонный потенциал возникает на границе волосковых клеток и эндолимфы улиткового протока и является изрецепторным потенциалом, предшествующим возникновению распространяющегося возбуждения в дендритах клеток спирального ганглия.
При действии звуков большой частоты и интенсивности различают суммационный потенциал, который представляет собой постоянный сдвиг потенциала эндолимфы улиткового протока относительно перилимфы барабанной лестницы.
Строение обонятельного анализатора. Периферическая часть обонятельного анализатора расположена в слизистой оболочке верхнего носового хода и противолежащей части носовой перегородки (рис. 70, А). Она представлена обонятельными и опорными клетками. Вокруг каждой опорной клетки расположено 9—10 обонятельных (рис. 70, Б). Обонятельные клетки покрыты волосками, которые представляют собой нити длиной 20—30 мкм. Они сгибаются и разгибаются со скоростью 20—50 раз в 1 мин. Внутри волосков расположены фибриллы, которые обычно заходят в утолщение — пуговку, имеющуюся на конце волоска. В теле обонятельной клетки и в ее периферическом отростке расположено большое количество микротрубочек диаметром 0,002 мкм, предполагают, что они осуществляют связь между различными органеллами клетки. Тело обонятельной клетки богато РНК, которая образует возле ядра плотные скопления. После воздействия паров пахучих


Рис. 70. Периферический отдел обонятельного анализатора:
д — схема строения носовой полости: 1 — нижний носовой ход; 2 — нижняя, 3 — средняя и 4 — верхняя носовые раковины; 5 — верхний носовой ход; Б — схема строения обонятельного эпителия: 1 — тело обонятельной клетки, 2 — опорная клетка; 3 — булава; 4 — микроворсинки; 5 — обонятельные нити.
веществ происходит их разрыхление и частичное исчезновение, что говорит о том, что функция обонятельных клеток сопровождается изменениями в распределении РНК и в ее количестве.
Обонятельная клетка имеет два отростка. Один из них через отверстия продырявленной пластинки решетчатой кости направляется в полость черепа к обонятельным луковицам, в которых возбуждение передается на расположенные там нейроны. Их волокна образуют обонятельные пути, которые подходят к различным отделам ствола мозга. Корковый отдел обонятельного анализатора находится в гиппокамповой извилине и в аммоновом роге.
Второй отросток обонятельной клетки имеет форму палочки шириной 1 мкм, длиной 20—30 мкм и заканчивается обонятельным пузырьком — булавой, диаметр которой 2 мкм. На обонятельном пузырьке расположено 9—16 ресничек.
Восприятие обонятельных раздражений. Обонятельные рецепторы обладают очень большой чувствительностью. Для возбуждения одной обонятельной клетки человека достаточно от 1 до 8 молекул пахучего вещества (бутилмеркаптана). Механизм восприятия запахов до настоящего времени еще не установлен. Предполагают, что обонятельные волоски являются как бы специализированными антеннами, которые активно участвуют в поиске и восприятии пахучих веществ. Относительно механизма восприятия существуют разные точки зрения. Так, Эймур (1962) считает, что на поверхности волосков обонятельных клеток расположены особые рецептивные участки в виде ямок, щелей определенного размера и определенным образом заряженных. Молекулы различных пахучих веществ имеют форму, размер и заряд, комплементарные различным участкам обонятельной клетки, и это обусловливает различение запахов.
Некоторые исследователи полагают, что обонятельный пигмент, имеющийся в обонятельной рецептивной зоне, также участвует в восприятии обонятельных раздражений, как пигмент сетчатки при восприятии зрительных раздражений. Согласно этим представлениям окрашенные формы пигмента содержат возбужденные электроны. Пахучие вещества, действуя на обонятельный пигмент, вызывают переход электронов на более низкий энергетический уровень, что сопровождается обесцвечиванием пигмента и освобождением энергии, которая затрачивается на возникновение импульсов.
Биопотенциалы возникают в булаве и распространяются далее по обонятельным путям до коры головного мозга.
Строение вкусового анализатора. Периферический отдел вкусового анализатора представлен вкусовыми луковицами, которые расположены главным образом в сосочках языка. Различают сосочки желобоватые, листовидные я грибовидные (рис. 71, А). В меньшем количестве вкусовые луковицы встречаются на мягком нёбе и задней стенке глотки. Вкусовая луковица имеет овальную форму и состоит из опорных и рецепторных вкусовых клеток (рис. 71, Б). Вкусовые клетки усеяны на своем конце микроворсинками, которые называют еще вкусовыми волосками. Длина микроворсинок — около 2 мкм, диаметр — около 0,2 мкм. Они выходят на поверхность языка через вкусовые поры.
На вкусовой клетке имеется большое число синапсов, которые образуют волокна барабанной струны и языкоглоточного нерва. Волокна барабанной струны (ветвь язычного нерва) подходят ко всем грибовидным сосочкам, а волокна языкоглоточного нерва — к желобоватым и листовидным. Корковый конец вкусового анализатора находится в гиппокампе, парагиппокамповой извилине и в нижней части заднецентральной извилины.
Вкусовые клетки непрерывно делятся и непрерывно гибнут. Особенно быстро происходит замещение клеток, расположенных в передней части языка, где они лежат более поверхностно. Замена клеток вкусовой почки сопровождается образованием новых синап-тических структур.
Восприятие вкусовых раздражителей. Микроворсинки вкусовых клеток являются образованиями, непосредственно воспринимающими вкусовой раздражитель. Мембранный потенциал вкусовых клеток колеблется от —30 до —50 мВ. При действии вкусовых раздражителей возникает рецепторный потенциал величиной от 15 до 40 мВ. Он представляет собой деполяризацию поверхности

Рис. 71. Периферический отдел вкусового анализатора:
А — сосочки языка: / — листовидный; 2 — грибовидный; 3 — желобоватый; Б — вкусовые клетки и опорные элементы: /— вкусовая пора; 2 — опорная клетка; 3 — рецепторная клетка; 4 — нервные волокна.
вкусовой клетки, которая является причиной возникновения в волокнах барабанной струны и языкоглоточного нерва генераторного потенциала, переходящего по достижении критического уровня в распространяющиеся импульсы. С рецепторной клетки возбуждение передается через синапс на нервное волокно с помощью ацетилхолина. Некоторые вещества, как например СаС12, хинин, соли тяжелых металлов, вызывают не первичную деполяризацию, а первичную гиперпрляризацию. Ее возникновение связано с осуществлением отрицательных отвергаемых реакций. Распространяющихся импульсов при этом не возникает.
Различные вкусовые клетки обладают разной чувствительностью к различным вкусовым веществам, которые делятся на четыре группы: кислое, соленое, сладкое, горькое. Каждая клетка отвечает всегда более чем на одно вкусовое вещество, иногда даже на все четыре, но наибольшей чувствительностью обладает к одному из них. Соответственно в зависимости от расположения клеток с особо высокой чувствительностью к тому или иному вкусовому раздражителю разные участки языка обладают также разной чувствительностью. Кончик языка наиболее чувствителен к сладкому, корень языка — к горькому.
Для вкусовых клеток характерны колебания порога раздражения и различный в разных условиях характер ответа на одни и те же раздражители. Их возбудимость зависит от постоянных влияний друг на друга, а также от состояния рецепторов пищеварительного тракта, обонятельных и др. В норме существует определенная «настройка» вкусовых рецепторов в соответствии с состоянием организма, в частности с состоянием сытости.
КОЖНЫЙ АНАЛИЗАТОР
Классификация и структура рецепторных образований кожного анализатора. К кожному анализатору относят совокупность анатомических образований, согласованной деятельностью которых определяются такие виды кожной чувствительности, как чувство давления, растяжения, прикосновения, вибрации, тепла, холода и боли.
Все рецепторные образования кожи в зависимости от их структуры делят на две группы: свободные и несвободные. Несвободные в свою очередь подразделяются на инкапсулированные и неинкапсулированные. Свободные нервные окончания представлены конечными разветвлениями дендритов сенсорных нейронов. Они теряют миелин, проникают между клетками эпителия и располагаются в эпидермисе и дерме. В некоторых случаях конечные разветвления осевого цилиндра окутывают измененные эпителиальные клетки, образуя осязательные мениски.
Несвободные нервные окончания состоят не только из ветвлений волокна, потерявшего миелин, но и из клеток глии. К несвободным инкапсулированным рецепторным образованиям кожи относятся пластинчатые тельца, или тельца Фатера-Пачини, осязательные


Рис. 72. Различные типы рецепторов кожи:
А — пластинчатое тельце Фатера-Пачини: 1 — наружная колба; 2 — концевой отдел нервного волокна; Б — осязательное мейснерово тельце; В — свободные нервные окончания; Г — осязательное тельце
Меркеля; Д — колба Краузе.
тельца, или тельца Мейснера, колбы Краузе и др. (рис. 72). Тельца Фатера-Пачини состоят из расположенной снаружи соединительнотканной капсулы и внутренней колбы. Последняя содержит измененные шванновские клетки. Во внутреннюю колбу входит, теряя при этом свою миелиновую оболочку, чувствительное нервное волокно. Тельца Мейснера состоят из тонкой соединительнотканной капсулы, внутри которой перпендикулярно к длинной оси тельца расположены глиальные клетки, накладывающиеся друг на друга. С поверхностью глиальных клеток контактируют разветвления нервного волокна, которые, входя в тельце, теряют миелин.
Колбы Краузе имеют сферическую форму, снаружи они одеты соединительнотканной капсулой. Нервные волокна, входящие внутрь колбы, сильно переплетаются.
Количество различного типа рецепторов, приходящихся на единицу поверхности, неодинаково. В среднем на 1 см2 кожной поверхности приходится 50 болевых, 25 тактильных, 12 Холодовых и 2 тепловые точки. Кожа различных участков тела имеет разное количество рецепторов и соответственно обладает неодинаковой чувствительностью. Особенно большое количество рецепторов расположено на поверхности губ, на кожной поверхности кончиков пальцев.
Функциональные свойства кожных рецепторов. Каждому виду чувствительности соответствуют особые рецепторные образования, которые делят на четыре группы: тактильные, тепловые, холодовые и. болевые.
Абсолютная специфичность, т. е. способность реагировать только на какой-то один вид раздражения, характерна лишь для некоторых рецепторных образований кожи. Многие из них реагируют на раздражители разной модальности. Возникновение различных ощущений зависит не только от того, какое рецепторное образование кожи подвергалось раздражению, но и от характера импульсации, идущей от этого рецептора в центральную нервную систему.
Способность к раздельному восприятию двух раздражений, наносимых на разные участки кожи, называют различительной чувствительностью.
Порог пространственного различения, под которым понимают наименьшее расстояние между двумя точками, раздражение которых воспринимается как раздельное, неодинаков для разных участков кожи. Так, на кончике языка он составляет 1 мм, на ладонной стороне ногтевых фаланг пальцев руки — 2 мм, на спине и груди — 40—70 мм.
Восприятие механических раздражений (прикосновение, давление, вибрации, растяжения) называют тактильной рецепцией. Она является одним из видов механорецепции, направленной на восприятие механической энергии раздражающего стимула. Механорецепция осуществляется рецепторами кожи, вестибулярного аппарата, мышц, сухожилий, стенок сосудов и др.
Функциональные свойства механорецепторов особенно детально были изучены на примере телец Фатера-Пачини. Эти рецепторные образования являются первично чувствующими. При действии механических раздражителей происходит растяжение рецепторной мембраны, увеличение ее проницаемости для ионов и возникновение генераторного потенциала.
По достижении критического уровня деполяризации в ближайшем перехвате Ранвье возникает ПД. Каждый рецепторный потенциал может дать начало 1—2 потенциалам действия.
Восприятие тепла и холода определяют как температурную рецепцию. Холодовые и тепловые рецепторы в определенном диапазоне температур обладают каким-то уровнем электрической активности. Для тепловых рецепторов теплокровных характерна импульсная активность в диапазоне температур от 20 до 50° С, а для Холодовых — от 10 до 41°С. Максимальная импульсация от тепловых рецепторов (2—15 имп/с) отмечается при 38—43°С, а от Холодовых—15—34°С. При быстром изменении температуры меняется частота импульсации. Холодовые рецепторы вызывают увеличение импульсации при охлаждении, тепловые—при согревании. У теплокровных изменение температуры на 0,2°С оказывается достаточным для изменения частоты импульсации в соответствующем афферентном проводнике.
Восприятие болевых раздражений осуществляется не только свободными нервными окончаниями, а при достаточно интенсивном раздражении и самыми различными рецепторами кожи.
Проводящие пути и корковый конец кожного анализатора. Возбуждение от рецепторов кожного анализатора направляется в центральную нервную систему по волокнам, имеющим различный диаметр. Волокна малого диаметра (со скоростью проведения возбуждения 30 м/с) осуществляют переключение на второй
нейрон в спинном мозге. Аксоны этих нейронов в составе передних и боковых восходящих путей направляются, частично перекрещиваясь, к зрительным буграм, где располагается третий нейрон пути кожной чувствительности. Отростки этих нейронов достигают соматосенсорной зоны пре- и постцентральной извилины коры.
Волокна более толстые (со скоростью проведения от 30 до 80 м/с) проходят без перерыва до продолговатого мозга, где и происходит переключение на второй нейрон. Там же осуществляется передача на второй нейрон возбуждения, идущего от рецепторов кожи головы. Аксоны нейронов продолговатого мозга полностью перекрещиваются на уровне продолговатого мозга и направляются к зрительным буграм. По аксонам нейронов зрительных бугров возбуждение передается в соматосенсорную область коры.
В зрительном бугре кожная поверхность головы и лица представлена в заднемедиальной зоне заднего вентрального ядра, а верхние и нижние конечности, туловище — в переднелатеральной его части. Имеется определенная организация и в расположении по вертикали нейронов, воспринимающих информацию от различных участков кожной поверхности. Выше всего расположены нейроны, воспринимающие информацию от кожной поверхности ног, несколько ниже — от туловища и еще ниже — от рук, шеи, головы. Такое же расположение характерно и для коркового отдела кожного анализатора (рис. 49). Нейроны, передающие информацию от кожной поверхности, делятся на моно-, ди- и полимодальные. Мономодальные нейроны выполняют функцию различения, а ди- и полимодальные— интегративную. Выделяют особые нейроны — «детекторы нового». Они отвечают очень кратковременной реакцией только на новые раздражения.

