2020-04-20
2020-04-20 726
726
В XX в., особенно начиная с 20‑х годов, широко развернулись экспериментальные исследования эмбрионального развития. Стремление преодолеть виталистическое истолкование эпигенеза, связанное, главным образом, с воззрением Дриша, вылилось в попытки представить формообразовательные процессы в онтогенезе как результат взаимного влияния частей развивающегося зародыша. Основателем этого направления в эмбриологии по справедливости считается немецкий эмбриолог Г. Шпеман, разработавший тонкие методы хирургического вмешательства в процессы эмбрионального развития.
Объектом исследования Шпемана, его учеников и многочисленных последователей были преимущественно зародыши амфибий на ранних стадиях развития. Используя простые инструменты – тонко отточенные скальпели, микропипетки, волосяные петли и стеклянные иглы, – эмбриологи шпемановской школы успешно осуществляли удаление определенных, точно локализованных участков зародыша и пересадки таких участков с одного места на другое того же или другого зародыша.
Если на стадии бластулы или ранней гаструлы пересадить участок будущей спинной эктодермы одного зародыша на брюшную или боковую сторону другого, то пересаженный участок развивается не так, как он развивался бы, будучи оставлен на месте. Этот участок дает начало не той или иной области головного или спинного мозга, а, ассимилируясь окружающей покровной эктодермой, принимает участие в образовании кожи. Другими словами, пересаженный участок эктодермы развивается не в соответствии с его происхождением, а в соответствии с местом, где он оказывается после операции. Равным образом, участок будущей покровной эктодермы, взятый с брюшной или боковой области бластулы или ранней гаструлы и пересаженный на спинную сторону, превращается не в кожу, которая должна была бы развиться на месте, откуда этот участок был изъят, а в часть нервной системы хозяина. Достоверность этих результатов обеспечивалась, в частности, применением гетеро‑ или ксенотрансплантации, т. е. пересадкой упомянутых участков от одного вида хвостатых амфибий другому, например, от зародыша гребенчатого тритона зародышу альпийского, или даже пересадкой в пределах разных отрядов амфибий, например, от зародыша бесхвостой амфибии – жерлянки зародышу хвостатой амфибии – тритону. Различия клеток трансплантата и клеток хозяина позволяли с уверенностью судить, какие части химерного зародыша развились из пересаженного материала и какие – из тканей хозяина под влиянием трансплантата.
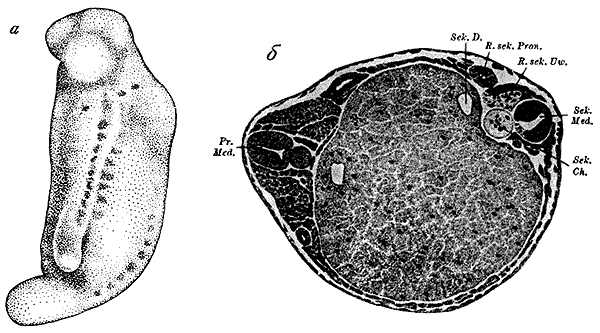
Индукция вторичного зародыша тритона (по Г. Шпеману и Г. Мангольд, 1924).
а – зародыш, на боку которого развился индуцированный зародыш со слуховыми пузырьками, медуллярной трубкой и двумя рядами сомитов; б – поперечный разрез через зародыш с первичными (слева) и вторичными (справа) индуцированными осевыми органами.
Результат получается иной, если описанные операции провести на стадии поздней гаструлы. В этом случае участок спинной эктодермы образует на брюшной или боковой стороне включенную в кожу часть центральной нервной системы, а участок брюшной или боковой эктодермы, пересаженной на спинную сторону, дает в центральной нервной системе начало тканям кожи. Иначе говоря, а этой стадии пересаженные на другое место участки зародыша развиваются уже не в соответствии с новым местоположением, а в соответствии с их происхождением. Из этих опытов был сделан вывод, что при переходе к стадии поздней гаструлы у зародыша утрачивается способность изменять направление развития под влиянием нового окружения и, наоборот, закрепляется способность развиваться в одном определенном направлении.