4.1.1. Общая характеристика нейронов
Нервная ткань превосходит все другие ткани по количеству генов, вовлеченных в развитие клеток, и по количеству видов информационной РНК. В мозге существует примерно 200 000 отличающихся друг от друга молекул
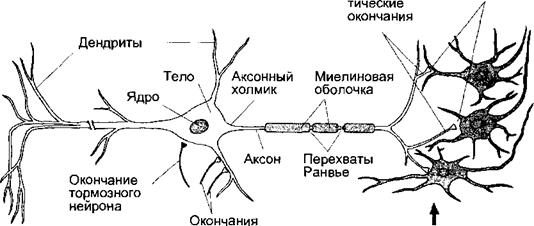
Дендриты — ветвящиеся отростки нейрона, представляющие область формирования входных сигналов. Аксон — отросток тела нейрона, служащий для проведения возбуждения от него, имеет постоянный диаметр (большинство аксонов в ЦНС окружены миелиновой оболочкой) и заканчивается пресинаптическими окончаниями. Аксонный холмик ~ конически расширенный участок начала аксона, представляющий наиболее возбудимую часть плазматической мембраны нейрона. Показаны окончания тормозного нейрона на теле клетки и возбуждающего нейрона на одном из дендритов.
иРНК, что в 10—20 раз больше, чем в печени или почках. Поэтому нервные клетки представлены многими популяциями нейронов, резко различающихся морфологическими признаками (строение тела, дендритов и аксонов) и синтезируемыми в них нейромедиаторами. В то же время активность любого нейрона, независимо от выполняемой функции, проявляется только в его возбуждении, т. е. генерации потенциалов действия, приводящей к выделению в синапсе нейромедиатора.
Выполняемая нейроном функция всецело определяется специфичностью его синаптических соединений с другими нейронами. Некоторые популяции нейронов образуют компактные ядра, предназначенные для переработки информации, которая передается из одного отдела мозга в другой. Другие популяции нейронов представлены небольшими скоплениями клеток, обладающих длинными разветвленными отростками, направленными сразу к нескольким регионам мозга для регуляции их активности.
Четыре морфологически различаемые области нейрона (тело, дендриты, аксон и пресинаптические окончания — рис. 4.1) отличаются друг от друга и в функциональном отношении. Тело нейрона является центром происходящих в ней обменных процессов. В теле нейрона находится ядро, где происходит переписывание генетической информации с ДНК на иРНК. Дендриты представляют собой разветвленные отростки нервной клетки, служащие для проведения нервных импульсов к телу нейрона. Степень разветвленности и общее количество дендритов коррелируют с количеством синапсов между нейроном и другими нервными клетками, способными повлиять на его активность и определяемыми как пресинаптические нейроны. Так, например, на дендритах мотонейронов спинного мозга в среднем существует около 8000 синапсов, а на дендритах клеток Пуркинье, находящихся в коре мозжечка, — 150 000. Если одни пресинаптические нейроны оказывают возбуждающее действие, а другие — тормозное, актив-
6 - 6095 ность постсинаптического нейрона будет определяться тем, какое из этих влияний преобладает.
Нейроны имеют один аксон, в большинстве случаев самый длинный отросток клетки, предназначенный для проведения нервных импульсов от ее тела. Обычно аксоны разделяются на несколько ветвей, или коллатералей, позволяющих нейрону активировать или тормозить с помощью своего медиатора сразу несколько клеток, с которыми он образует синапсы. В плазматической мембране аксона содержится большое количество потенциалзависимых каналов для ионов натрия, необходимых для генерации и проведения потенциалов действия. Наибольшая плотность таких каналов характерна для аксонного холмика — суживающегося начального участка аксона, отходящего от клеточного тела. Это наиболее возбудимый участок мембраны центральных нейронов, где легче всего достигается критический уровень деполяризации. Именно поэтому потенциалы действия обычно возникают в аксонном холмике, представляющим собой наиболее чувствительную к возбуждению область клетки.
Пресинаптические окончания аксонов свободны от миелинового покрытия и предназначены для хранения и выделения медиаторов в химических синапсах. К выделению медиаторов приводит входящий ток ионов кальция через быстро инактивирующиеся каналы, плотность которых в пресинап- тических окончаниях высока. Аксоны возбуждающих нейронов в большинстве случаев образуют синапсы с дендритами и реже с телом постсинаптических нейронов. Аксоны тормозных нейронов чаще всего образуют синапсы на теле постсинаптического нейрона, их действие создает препятствие проведению возбуждения от дендритов к аксонному холмику. Аксо- аксоналъные синапсы, которые могут быть как возбуждающими, так и тормозными, не влияют непосредственно на зону возникновения электрического сигнала. Их функция заключается в модуляции выходной активности постсинаптического нейрона, т. е. в увеличении или уменьшении количества выделяемого в окончаниях его аксона медиатора.
4.1.2. Функциональная модель нейрона
Функция нейрона заключается в его способности возбуждаться и, действуя через синапсы на другие клетки, генерировать возбуждение в них, что приводит к распространению этого процесса от одного нейрона к другому. Возбуждение тормозного нейрона сопровождается подавлением процесса возбуждения в клетках, на которые он действует. Процесс возбуждения включает в себя возникновение нескольких типов электрических сигналов, образование и распространение которых происходит по единым правилам во всех нервных клетках: сенсорных, моторных, вставочных и нейросекреторных. Это позволяет рассмотреть закономерности возникновения и распространения электрических сигналов на основе единой функциональной модели нейрона, пригодной для характеристики любого типа нервных клеток.
Функциональная модель нейрона предусматривает выделение в нем четырех функциональных областей, каждая из которых предназначена для формирования одной из четырех разновидностей сигналов, характеризующих процесс возбуждения: 1) входного (постсинаптический и рецепторный потенциалы), 2) объединенного (потенциал действия), 3) проводящегося и 4) выходного (выделение медиатора) (рис. 4.2).
4.1.2.1. Входные сигналы
Входные сигналы представляют собой изменения величины мембранного потенциала в результате действия химических медиаторов на мембраны постсинаптических нейронов (постсинаптические потенциалы), а в чувствительных окончаниях сенсорных нейронов они возникают в ответ на действие адекватных раздражителей (рецепторный потенциал). В возбуждающих синапсах ЦНС входной сигнал представляет собой деполяризацию постсинаптической мембраны на 0,2—0,3 мВ, в тормозных синапсах — столь же незначительную гиперполяризацию, соответственно этому различают возбуждающие постсинаптические потенциалы (ВПСП) и тормозные постсинаптические потенциалы (ТПСП).
Постсинаптические потенциалы могут распространяться на очень малое расстояние от места возникновения и в связи с этим определяются как местные, или локальные, потенциалы. Их амплитуда пропорциональна количеству молекул нейромедиатора, связавшегося с постсинаптическими рецепторами, такая зависимость является градуальной, а поэтому и входные сигналы градуальны. Назначение возбуждающих входных сигналов состоит в том, чтобы деполяризовать мембрану нейрона до критической величины, достаточной для генерации потенциалов действия, а тормозные входные сигналы предназначены для того, чтобы этому препятствовать.
Одиночные ВПСП электротонически, т. е. пассивно, распространяются от места возникновения (дендриты или тело нейрона) к аксонному холмику, служащему интегративной зоной или местом генерации потенциалов действия. Изначально малая амплитуда одиночных ВПСП при пассивном проведении становится еще меньше (рис. 4.3), поэтому для достижения



| Проведение объединенного сигнала
| |
Различные по выполняемой функции нейроны (сенсорные, моторные, интернейроны, нейросекретирующие клетки) имеют четыре функционально сопоставимые области: входную, интегративную, проводящую и выходную. Эти области предназначены для возникновения или проведения электрических сигналов, среди которых различают: входной (рецепторный или постсииаптический потенциалы), интегративный (объединенный сигнал или потенциал действия), проводящийся сигнал и выходной сигнал.
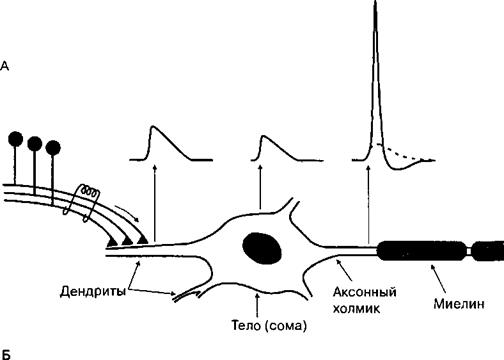
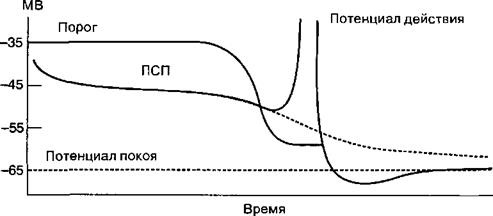
Рис. 4.3. Уменьшение амплитуды постсинаптического потенциала (ПСП) при распространении его по мембране клетки
А. В результате возбуждения окончаний пресинаптических нейронов и выделения из них нейромедиатора на дендрите постсинаптического нейрона образуется возбуждающий постсинаптический потенциал (ВПСП). Амплитуда ВПСП уменьшается по мере его распространения от дендрита к телу нейрона.
Б. Величина потенциала покоя постсинаптического нейрона составляет —65 мВ, порог возбуждения (уровень критической деполяризации) на теле нейрона соответствует —35 мВ, а в области аксонного холмика — около —50 мВ. Несмотря на уменьшение амплитуды распространяющегося по мембране ВПСП, в области аксонного холмика деполяризующий сдвиг достигает критического значения.
критической деполяризации мембраны необходима пространственная или последовательная (временная) суммация ВПСП.
Если постсинаптические потенциалы возникнут одновременно в разных синапсах на дендритах и теле нейрона, то их общая сумма может оказаться достаточной для деполяризации мембраны интегративной зоны до критического уровня. Эта разновидность суммации называется пространственной, она возникает при одновременной возбуждающей активности группы нейронов, аксоны которых конвергируют к одной общей постсинаптической клетке (рис. 4.4). Средняя величина мембранного потенциала покоя у

| Б. Последовательная суммация
| |
А. Пространственная суммация возникает при одновременном возбуждении нейронов, конвергирующих к общей постсинаптической клетке.
Б. Последовательная суммация происходит вследствие ритмического возбуждения пресинапти- ческого нейрона с достаточно высокой частотой генерации потенциалов действия (через каждые 10 мс).
центральных нейронов составляет—65 мВ, а среднее значение критической деполяризации —55 мВ, исходя из этого, для возбуждения постсинаптического нейрона необходима сумма ВПСП, превышающая 10 мВ. Деполяризующий сдвиг, превышающий это значение, приводит к возбуждению постсинаптического нейрона.
Последовательная или временная суммация ВПСП происходит вследствие продолжительного возбуждения пресинаптического нейрона, генерирующего потенциалы действия, которые следуют друг за другом через короткие промежутки времени. Если 50—100 нервных импульсов достигнут пресинаптического окончания непосредственно друг за другом, то общего количества выделившегося медиатора окажется достаточно для получения амплитуды ВПСП, превышающей уровень критической деполяризации, что вызовет возбуждение постсинаптического нейрона. В центральных синапсах ЦНС оба вида суммации, пространственная и последовательная, обычно происходят одновременно, что способствует возбуждению постсинаптических нейронов.
На дендритах, но чаще на теле клетки существуют тормозные синапсы, расположенные между возбуждающими синапсами и интегративной зоной. Их функция состоит в том, чтобы помешать деполяризующим ВПСП достичь интегративной зоны. В тормозных синапсах активируются хемозави- симые каналы для ионов калия или хлора, что ведет к гиперполяризации мембраны, представляющей тормозной постсинаптический потенциал. Если на пути распространения деполяризующих ВПСП окажутся гиперполя- ризованные действием тормозных медиаторов участки мембраны, то суммарная амплитуда ВПСП не сможет достичь значения критической деполяризации, необходимой для возникновения потенциала действия
| Раздельное действие возбуждающих и тормозного нейронов
Е кр.
| |
| ВПСП после действия тормозного нейрона
| |
 2020-05-25
2020-05-25 439
439