 2020-06-08
2020-06-08 267
2671 - мембранатектория;
2 — внутренние чувствительные клетки;
3 — наружные чувствительные клетки;
4 — нервные волокна;
5 — клетки Хенсена;
6 — клетки Клаудиса;
7 — базальная мембрана;
8 — спиральный ганглий.
92
окно, верхнюю и нижнюю лестницы, соединенные геликотремой, заканчивается круглым окном. Верхний и нижний каналы улитки заполнены перилимфой, а средний — эндолимфой. Перилимфа напоминает плазму крови и цереброспинальную жидкость, в которой преобладает содержание ионов натрия. Эндолимфа отличается от перилимфы высокой концентрацией ионов калия, приближаясь по химическому составу к внутриклеточной жидкости.
Основная мембрана состоит из эластических волокон, слабо натянутых между костным спиральным гребешком и наружной стенкой улитки, что создает условия для колебательных движений волокон базилярной мембраны. На основной мембране в средней лестнице расположен звуковоспринимающий рецепторный аппарат — кортиев орган. Кортнев орган состоит из четырех рядов волосковых клеток. Поверх волосков, или волосковых клеток, омываемых эндолимфой, лежит, соприкасаясь с ними, покровная, или текториальная мембрана.
Проведение звуковых колебаний в улитке. Звуковая волна, воздействуя на систему слуховых косточек среднего уха, приводит в колебательное движение мембрану овального окна, которая, прогибаясь, вызывает волнообразные перемещения перилимфы верхнего и нижнего каналов, которые постепенно затухают по направлению к вершине улитки. Колебания перилимфы передаются на вестибулярную мембрану, а также на полость среднего канала, приводя в движение эндолимфу и базилярную мембрану. Установлено, что при действии на ухо звуков низкой частоты (до 1000 Гц) происходит смещение базилярной мембраны на всем ее протяжении от основания до верхушки улитки. При увеличении частоты звукового сигнала происходит перемещение укороченного по длине колеблющегося столба жидкости ближе к овальному окну и наиболее жесткому и упругому участку базилярной мембраны. Деформируясь, базилярная мембрана смещает волоски волосковых клеток относительно текториальной мембраны. ЕГ результате такого смещения возникает электрический разряд волосковых клеток. Существует прямая зависимость между амплитудой смещения основной мембраны и количеством вовлекаемых в процесс возбуждения нейронов слуховой коры. Электрофизиологические исследования показали, что средний канал улитки имеет положительный заряд относительно верхнего и нижнего каналов. Это — эндокохлеарный потенциал улитки. Он обусловлен определенным уровнем окислительно-восстановительных процессов в каналах улитки. Разрушение сосудистой оболочки и гипоксия приводят к его исчезновению. Эндокохлеарный потенциал создает критический уровень поляризации волосковых клеток, поэтому незначительное механическое воздействие приводит к возникновению возбуждения в волосковых клетках. В этом, видимо, и состоит основное функциональное значение. Различают три вида электрических реакций во внутреннем ухе: 1) микрофонный эффект, 2) суммационный потенциал, 3) потенциал действия слухового нерва.
Впервые микрофонный эффект улитки был получен Е. Уивером и С. Бреем в 1930 г. В эксперименте на кошках было показано, что если в улитку ввести электроды, соединенные с усилителем и громкоговорителем, расположенным в другом помещении, а затем на ухо кошке произносить различные слова, то экспериментатор, находясь у громкоговорителя в другом помещении, может услышать эти же слова. Микрофонный эффект улитки возникает в ответ на смещение текториальной мембраны относительно волосковых клеток, по форме и частоте напоминая форму звуковых колебаний. Происхождение микрофонного эффекта связывают с механохимическими преобразованиями в волосковых клетках корти- ева органа, повреждение которого приводит к исчезновению микрофонного эффекта. Высокоамплитудные потенциалы отводят от той части улитки, резонансная частота которой одинакова с частотой действующих на ухо звуковых колебаний. Микрофонный потенциал регистрируется еще некоторое время после смерти животного, но его частотные и амплитудные характеристики убывают.
Местом возникновения микрофонного потенциала является область корешков волосков волосковых клеток. Звуковые колебания, действующие на внутреннее ухо, накладывают возникающий микрофонный эффект на эндокохлеарный потенциал, вызывая его модуляцию.
Суммарный потенциал отличается от микрофонного потенциала тем, что отражает не форму звуковой волны, а ее огибающую и возникает при действии на ухо высокочастотных звуков.
Потенциал действия слухового нерва генерируется в результате электрического возбуждения, возникающего в волосковых клетках.
Электронно-микроскопические исследования показали наличие синаптических контактов между волосковыми клетками и нервными окончаниями. Предполагают химический способ передачи возбуждения с волосковых клеток на волокна слухового нерва. Потенциал действия в нервных окончаниях регистрируется через 0,5—1,0 мс после возникновения микрофонного эффекта, что также говорит в пользу синаптической передачи возбуждения.
Восприятие звука различной частоты. В настоящее время распространена «теория места». Предполагают, что волосковые клетки, расположенные на базилярной мембране в различных участках улитки, обладают разной лабильностью, что оказывает влияние на восприятие звуков высокой и низкой частоты (настройку волосковых клеток на звуки различной частоты).
Проводящие пути и центры слухового анализатора. Нейроны первого порядка слухового пути входят в состав спирального ганглия улитки. Центральные отростки клеток спирального ганглия образуют слуховой, или кохлеарный, нерв. Периферические отростки этих же клеток идут по направлению к кортиеву органу. Кохлеарный нерв, являясь ветвью VIII пары черепно-мозговых нервов, проходит в продолговатый мозг и заканчивается на клетках кохлеарных ядер (нейроны второго порядка). Все три ядра составляют так называемый кохлеарный комплекс. Улитка представлена в ядрах кохлеарного комплекса таким образом, что волокна, идущие от верхушки улитки, оканчиваются в вентролатеральном отделе комплекса; идущие от основания улитки — в его дорсомедиальных частях. От нейронов кохлеарного комплекса начинается восходящий слуховой путь, который делится на ипси и, более мощный, контрлатеральный пучок волокон. Контрлатеральные волокна оканчиваются на клетках верхней оливы. Аксоны нейронов верхней оливы вместе с непереключенными волокнами проходят в составе латеральной петли. Одна часть волокон латеральной петли достигает ядер нижних бугров четверохолмия, в которых представлены нейроны Ш, IV, V порядков. Другая часть волокон латеральной петли проходит, не переключаясь, во внутреннее коленчатое тело зрительного бугра данной стороны, которое является последним переключательным звеном восходящего слухового пути. От внутренних, или медиальных, коленчатых тел волокна достигают клеток слуховой коры, заканчиваясь в верхней части височной доли мозга (поля 41 и 42 по Бродману).
Нисходящие пути слухового анализатора начинаются от клеток слуховой коры, переключаясь последовательно в медиальных коленчатых телах зрительного бугра, задних буграх четверохолмия, верхнеоливарном комплексе. Затем входят в кохлеарный нерв, достигая волосковых клеток кортиева органа.
Переработка информации в центрах. Функция отдельных частей проводящей системы слухового анализатора состоит в следующем. Клетки кортиева органа кодируют информацию. Нижние бугры четверохолмия отвечают за воспроизведение ориентировочного рефлекса на звуковое раздражение (поворот головы в сторону источника звука). Слуховая кора принимает активное участие в обработке информации, связанной с анализом коротких звуковых сигналов, с процессом дифференцировки звуков, фиксацией начального момента звука, различения его деятельности. Слуховая кора ответственна за создание комплексного представления о звуковом сигнале, поступающем в оба уха раздельно, а также за пространственную локализацию звуковых сигналов. Нейроны, участвующие в обработке информации, идущей от слуховых рецепторов, специализируются по выделению (детектированию) соответствующих признаков. Особенно эта дифференцировка присуща нейронам слуховой коры, расположенным в верхней височной извилине. Здесь имеются колонки, которые анализируют поступающую информацию. Среди нейронов слуховой коры выделяют так называемые простые нейроны, функции которых — вычленение информации о чистых звуках. Есть нейроны, которые возбуждаются только на определенную последовательность звуков или на определенную амплитудную их модуляцию. Есть нейроны, которые позволяют определить
94
направление звука. Т. о. происходит сложнейший анализ звукового сигнала. Однако представление о мелодии возникает в ассоциативных участках коры, в которых осуществляется сложнейший анализ поступающей информации на основе информации, хранящейся в памяти. Именно в ассоциативных участках коры с помощью специализированных нейронов мы способны извлечь всю информацию, поступающую от соответствующих рецепторов.
Длительное воздействие надлорогового звука вызывает утомление слухового анализа - тора, которое выражается в значительном снижении слуховой чувствительности и замедленном ее восстановлении. Так, у лиц, работающих в шумных цехах, вначале возникает утомление слуховой системы, а затем может развиться тугоухость, сопровождающаяся изменениями в волосковых клетках кортиева органа.
В механизме слуховой адаптации принимают участие- как периферические, так и центральные отделы слухового анализатора. Ослабление рассмотренного выше рефлекса мышц среднего уха лежит в основе адаптивных механизмов периферического отдела слухового анализатора. Значительную долю участия в механизме адаптации принимают центральные отделы слухового анализатора. И, в частности, было показано, что слуховая адаптация регулируется ретикулярными структурами ствола мозга и задним гипоталамусом.
Слуховая ориентация в пространстве происходит двумя путями. В первом случае определяется местоположение самого звучащего объекта (первичная локализация), во втором — происходит восприятие отраженных от различных объектов звуковых волн. Таким объектом может быть животное или человек. Это так называемая вторичная локализация звука, или эхолокация. При помощи эхолокации ориентируются в пространстве некоторые животные (дельфины, летучие мыши), а также люди, потерявшие зрение, или с нормальным зрением, но в условиях темноты. Пространственное восприятие звука возможно при наличии бинаурального слуха: способности определить местонахождения источника звука одновременно правым и левым ухом. При односторонней глухоте определение местоположения источника звука одним ухом облегчается поворотом головы в сторону звучащего источника, локализация которого в пространстве происходит путем сопоставления рисунка возбуждения в различных частях слуховой системы. Корковый конец слухового анализатора играет существенную роль в локализации источника звука в пространстве. Так, двустороннее удаление слуховой коры приводит к значительным нарушениям пространственного слуха.
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Глазное яблоко. Важнейшая информация поступает из внешней среды через зрительный анализатор. Периферический отдел зрительного анализатора особенно сложен. Он представлен глазным яблоком. Последнее является системой, преломляющей световые лучи. К преломляющим средам относятся роговица, жидкость передней камеры глаза, хрусталик и стекловидное тело. Радужная оболочка, как диафрагма в фотоаппарате, регулирует поток света. Заложенные в ней циркулярные мышцы получают парасимпатическую иннервацию, радиальные — симпатическую. При повышении тонуса парасимпатического отдела нервной системы величина зрачка уменьшается, при повышении тонуса симпатического отдела — увеличивается.
Хрусталик имеет форму двояковыпуклой линзы. Основная функция хрусталика состоит в преломлении проходящих через него лучей светд и фокусировки изображения на сетчатке. Преломляющая сила хрусталика непостоянна и, благодаря тому, что он может принимать более выпуклую форму, колеблется от 19 до 33 диоптрий. Изменение формы хрусталика (аккомодация) достигается при сокращении или расслаблении цилиарной мышцы, которая прикрепляется к капсуле хрусталика посредством цинновых связок.
Предполагается, что механизм аккомодации обеспечивается подкорковыми и корковыми зрительными центрами. Эти образования регулируют тонус цилиарной мышцы. В связи с тем, что хрусталик не является идеальной линзой, лучи света, проходящие через его периферическую часть, преломляются сильнее, в результате чего возникает искажение изо-
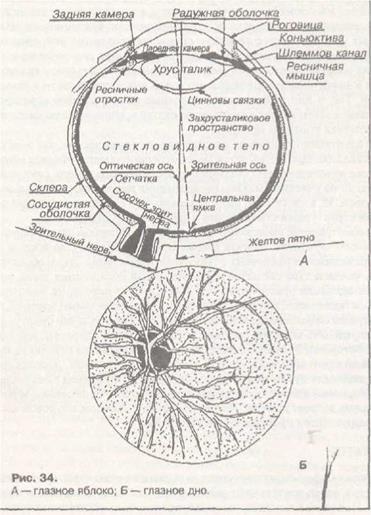
бражения — сферическая аберрация. Свет различной длины волн также преломляется хрусталиком неодинаково, и возникает хроматическая аберрация. Астигматизм — это дефект светопреломляющих сред глаза, связанный с неодинаковой кривизной их преломляющих поверхностей. Так, если кривизна поверхности роговицы в вертикальном сечении больше, чем в горизонтальном, изображение на сетчатке не будет четким независимо от расстояния до предмета. Роговица будет иметь как бы два главных фокуса: один — для вертикального сечения, другой — для горизонтального. Поэтому лучи света, проходящие через астигматический глаз, будут фокусироваться в разных плоскостях: если горизонтальные линии предмета будут сфокусированы на сетчатке, то вертикальные — впереди нее.
В течение жизни
хрусталик постепенно утрачивает свои основные свойства — прозрачность и эластичность. Сила аккомодации уменьшается, и точка ближайшего ясного видения отодвигается вдаль. Развивается старческая дальнозоркость, или пресбиопия.
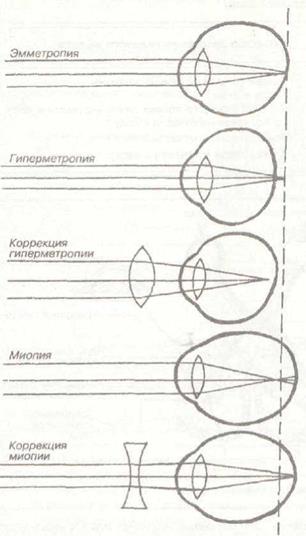
Необходимыми питательными веществами хрусталик обеспечивается за счет диффузии их из окружающей его жидкости. В связи с этим внутренние его слои находятся, с точки зрения обмена веществ, в наиболее неблагоприятных условиях. Может возникнуть постепенная дегенерация внутренних слоев хрусталика, что вызывает его помутнение и потерю эластичности. Нормальное зрение называется эмметропическим.
В связи с анатомическими дефектами глазного яблока (удлиненный или короткий глаз) возникают нарушения рефракции, что характеризуется близорукостью или дальнозоркостью. Миопия, или близорукость, возникает в том случае, когда при расслабленной аккомодации главный фокус оптической системы глаза располагается впереди сетчатки. Явление миопии характерно для удлиненного глаза. Гиперметропия, или дальнозоркость, присуща укороченному глазу.,В этом случае зона четкого изображения отодвигается эа сетчатку. При гиперметропии возможна самостоятельная коррекция путем напряжения аккомодации. Если это напряжение невелико, то малая степень дальнозоркости ничем себя не прояв-
96
 |
ляет. При большей степени гиперметропии необходима коррекция рефракции. При данных видах нарушения рефракции глаза сила аккомодации остается, как правило, нормальной, в отличие от пресбиопии, когда уменьшение широты аккомодации приводит к развитию дальнозоркости.
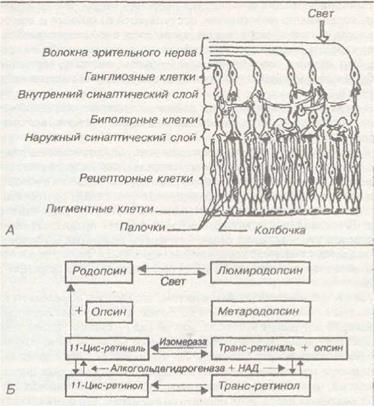
Проводящие пути зрительного анализатора. Три первых нейрона зрительных путей заложены в сетчатке: клетки с окончанием в виде палочек и колбочек, которые передают импульсы биополярным клеткам, а те — ганглиозным клеткам. Аксоны ганглиозных клеток составляют зрительный нерв. В области турецкого седла происходит частичный перекрест зрительного нерва и формируются два зрительных тракта. Каждый несет в себе волокна правого и левого глаза. Они заканчиваются в подкорковых центрах: латеральных коленчатых телах, верхних буграх четверохолмия и подушке зрительного бугра. Отсюда волокна отправляются в затылочную облатлвкоры.
Обработка информации в центрах. Обработка информации в этом анализаторе начинается на периферии — непосредственно на сетчатке. Собственно
Рис. 35. Схематическое представление о видах фоторецептор (палочка или
рефракции и их компенсации. колбочка) устроен таким образом, что
ствующейдлинысветавнем происходит изменение: хромофорная группа зрительного пигмента (цис-ретиналь) поглощает квант света и под влиянием избыточной энергии переходит в другую форму (транс-ретиналь), это приводит к тому, что ретиналь отщепляется от белка-носителя (опсина); одновременно происходит высвобождение молскулы-псрсносчика сигнала, скорее всего — ионов кальция. Эти ионы (или молекулы-переносчики) подходят к мембране рецептора и закрывают натриевые каналы. В результате происходит гиперполяризация (генерация рецепторного потенциала). Это единственное исключение из правила, когда рецепторный потенциал является гиперполяризующим, а не деполяризующим (как во всех других рецепторных образованиях). Что же происходит дальше? Рецепторная клетка контактирует с биполярной клеткой, которая в условиях темноты находится в по-
7. Физиология человека
Таблица 3. Движение глаз. Функция глазных мышц
 Характер движения глазного яблока
Характер движения глазного яблока
 |
|
|
 | |||
 | |||
|
|
 | ||||||
 | ||||||
 | ||||||
 | ||||||
Поворот глазного яблока кнаружи Тоже кнутри Поворот глазного яблока книзу и отчасти к носу То же, кверху и отчасти к носу То же, кверху и отчасти к виску То же, книзу и отчасти к виску
стоянном гиперполяризующем состоянии. Это состояние возникает под влиянием непрерывно выделяемого медиатора из фоторецептора. Медиатор вызывает гиперполяризацию биполярной клетки. Когда происходит взаимодействие фоторецептора с квантом света и (как отмечалось выше) гиперполяризация, то это вызывает уменьшение образования медиатора, поэтому снижается способность медиатора гиперполяризовать биполярную клетку. В связи с этим в условиях освещения биполярная клетка деполяризуется. Это приводит к тому, что на другой стороне биполярной клетки выделяется второй медиатор, который на мембране ганглиозной клетки вызывает деполяризацию (генераторный потенциал), что приводит к активации зрительного нерва. Поток ПД идет в латеральное коленчатое тело и в передние (верхние) бугры четверохолмия, а оттуда (по двум различным путям) достигает зрительной коры (поля 17, 18 и 19), расположенной в затылочной доле. Оттуда информация поступает в лобные и теменные доли — ассоциативные зоны коры, где формируется ответ на вопрос «Что такое?».
Как же происходит анализ зрительной информации?
Прежде всего — за счет организации рецептивных зон и соответствующих нейронов. Все рецепторные поля (зоны) имеют концентрический вид (окружности разного диаметра).
98
|
 нейронов), что они позволяют, во-первых, оценить — в какой части рецептивного поля находится световой луч, а во- вторых, — что происходит с сетчаткой: освещается она или нет. Часть рецептивных полей устроена таким образом, что их нейроны (назовем их оп-нейроны) возбуждаются в том случае, если освещается центр этого поля, а периферия не возбуждается. Второй вариант организации рецептивного поля заключается в том, что нейрон (off-нейрон) будет возбуждаться только в том случае, если освещается периферия. В сетчатке имеются ганглиозные клетки, которые реагируют возбуждением, например, на освещение центра, либо постоянным возбуждением (генерируют непрерывно ПД) — это медленно адаптирующие нейроны, либо только на момент включения (быстроадап- тирующие нейроны), другие нейроны возбуждаются только в случае, если стимул «движется» — перемещается по сетчатке. Благодаря сложной организации и специализации нейронов сетчатки уже на этом уровне происходит определение таких сложных качеств светового стимула, как освещенность, цвет, форма, движение сигнала.
нейронов), что они позволяют, во-первых, оценить — в какой части рецептивного поля находится световой луч, а во- вторых, — что происходит с сетчаткой: освещается она или нет. Часть рецептивных полей устроена таким образом, что их нейроны (назовем их оп-нейроны) возбуждаются в том случае, если освещается центр этого поля, а периферия не возбуждается. Второй вариант организации рецептивного поля заключается в том, что нейрон (off-нейрон) будет возбуждаться только в том случае, если освещается периферия. В сетчатке имеются ганглиозные клетки, которые реагируют возбуждением, например, на освещение центра, либо постоянным возбуждением (генерируют непрерывно ПД) — это медленно адаптирующие нейроны, либо только на момент включения (быстроадап- тирующие нейроны), другие нейроны возбуждаются только в случае, если стимул «движется» — перемещается по сетчатке. Благодаря сложной организации и специализации нейронов сетчатки уже на этом уровне происходит определение таких сложных качеств светового стимула, как освещенность, цвет, форма, движение сигнала.
В остальных передаточных станциях: передние или верхние бугры четверохолмия, латеральное коленчатое тело как специфическое ядро таламуса, предназначенное для переработки зрительной информации, происходит вычленение этой информации и выявление новых качеств, недоступных «примитивной» сетчатке. С этой целью за счет явления конвергенции и дивергенции создаются более сложные рецептивные поля, а также появляются более «обученные», более «смышленые» нейроны, которые возбуждаются на особые сигналы, например, на стимул, траектория движения которого имеет волнообразный характер. Уже на уровне передних бугров четверохолмия имеет место сохранение топического расположения рецепторов на сетчатке, а также наличие колонок — вертикальных скоплений нейронов, предназначенных для расчленения информации, поступающей от данной области сетчатки, на отдельные составляющие. Например, в латеральном коленчатом теле удалось выявить нейроны, отвечающие за контраст или реагирующие на цвет.
Основной анализ зрительной информации совершается нейронами коры. В первичном проекционном поле 17 происходит анализ информации, поступающей из правого и левого глаз. Как и в других зонах коры, в этой области анализ проводится с участием колонок. Имеются глазодоминантные колонки, анализирующие информациюЛ идущую либо из правого глаза, либо из левого. Эти колонки соседствуют друг с другом, поэтому, вероятно, между ними происходит обмен информацией, и это позволяет видеть двумя глазами один предмет(бинокулярное зрение).
Среди нейронов колонок коры различают «простые» нейроны, задача которых выявить контраст, наличие движущегося стимула, т. е. точно такие же задачи, как у нейронов сетчатки (но здесь рецептивные поля уже обобщены).
Имеются «сложные» и «сверхсложные» нейроны, которые возбуждаются при наличии определенных условий. Например, при движении светового сигнала слева направо или снизу вверх и т. п. Вей информация от нейронов проекционного поля коры передается в ассоциативные Зоны коры; где уже за счет более «обученных» нейронов происходит окончательное формирование образа. Здесь имеются нейроны, обученные узнавать все буквы нашего алфавита, слова, лица и т. д. И когда соответствующий нейрон «узнает» предназначенное ему узнавать, то он возбуждается, что и является физиологическим механизмом восприятия. Ассоциативные зоны находятся в теменной коре, в лобных извилинах. Если у больного повреждены теменные ассоциативные участки, то он перестает узнавать. Это приводит к развитию зрительной агнозии.
Теория цветоощущения. Все исследователи сходятся на том, что цвет мы определяем на основе рецепции световой волны с помощью трех видов колбочек: один вид наиболее чувствителен к длине волны, дающий ощущение красного, другой вид — синего (фиолетового), а третий вид колбочек дает ощущение желтого (принятое ранее представление о наличии «зеленоузнающих» колбочек подвергнуто ревизии). Но что дальше? Еще в прошлом веке физиолог Э. Геринг выдвинул представление о так называемых оппонентных цветах (красный-зеленый, синий-желтый, черный-белый). Оказалось, что его теория хорошо объясняет способность человека различать цвета, если принять во внимание, что функцию различения цветов выполняет нейрон, рецептивное поле которого устроено следующим образом: в центре находятся колбочки, воспринимающие, к примеру, красный цвет, а на периферии — колбочки, которые нечувствительны к нему: когда луч красного цвета возбуждает колбочки, находящиеся в центре рецептивного поля, нейрон возбуждается и, в конечном итоге, у нас возникает ощущение красного. Если свет в основном поглощается колбочками на периферии рецептивного поля, то нейрон этот не возбуждается, и в итоге у нас создается ощущение зеленого (но при условии существования второго варианта рецептивного поля: в центре находятся колбочки, воспринимающие зеленый цвет, тогда возбуждение этих нейронов даст ощущение зеленого, если же кванты воспринимаются в основном колбочками периферии, то нейрон не возбуждается, и это вместе с другим типом объединения рецепторов дает ощущение красного).
Синий и желтый цвет возникает в рецептивных полях, где в одном случае центр представлен колбочками, воспринимающими желтый цвет, а периферия — синий, и наоборот, рецептивные поля, где в центре концентрируются колбочки, воспринимающие синий цвет, а на периферии — желтый.
Черный-белый цвета образуются в результате такой организации, когда центр воспринимает все цвета (т. е. колбочки чувствительны ко всем цветам — это дает ощущение белого цвета), а периферия — нечувствительна к свету. Таким образом, любой цвет может быть представлен совокупностью нейронов, воспринимающих основные цвета. В результате многочисленного объединения нейронов на более высоком уровне (латеральное коленчатое тело, кора) возникает ощущение всех оттенков цвета.
При отсутствии какого-либо вида колбочек возникает аномалия цветовосприятия.
Таким образом, трехкомпонентная теория цветовосприятия (колбочки трех видов) хорошо согласуется с оппонентной теорией.
100
ОБОНЯТЕЛЬНЫЙ АНАЛИЗАТОР
Рецепторы обонятельного анализатора заложены в слизистой носа в области верхней носовой раковины. Они представляют собой чувствительные волосковые клетки, располагающиеся среди опорных клеток, включенных в эпителий. Нервные волокна, отходящие от чувствительных клеток, составляют обонятельные нервы, заканчивающиеся обонятельными луковицами. Последние имеют очень сложное строение — складываются из шести слоев специализированных нейронов, в которых происходит первичная переработка информации. Аксоны этих клеток направляются в подкорковые центры, нейроны которых дают аксоны, поступающие в корковые центры — в области ункус гиплокампа (предположительно). Различные нейроны вкусовых луковиц, как показали электрофизиологические исследования, по-разному реагируют на пахучие вещества разного вида (в определенной мере специализированы). Классификацию запахов не считают исчерпывающей. Имеет хождение подразделение запахов на цветочный, кислый, горелый, гнилостный. Каждый из них имеет огромное число разнообразных оттенков, воспринимаемых не только обонятельными окончаниями, но также вкусовыми, тактильными и другими рецепторами. Минимальные количества пахучего вещества, вызывающие ощущение запаха, называются пороговыми. Их можно определить с помощью прибора — ольфактометра., Одной из наиболее распространенных теорий восприятия запаха является стереохимическая. Предполагается, что на мембране обонятельных клеток имеются участки определенной конфигурации, адсорбирующие молекулы соответствующей формы. При взаимодействии молекулы с рецептором в нервном окончании генерируется потенциал, передающийся по волокнам в центры. Применяя запахи разного типа, исследователи получали разные электрофизиологические паттерны на запахи: камфорный, мускусный, цветочный, мятный, эфирный и т. д. Роль обонятельного анализатора у человека по сравнению с животными несоизмеримо мала.
ВКУСОВОЙ АНАЛИЗАТОР
Вкусовые рецепторы заложены в сосочках языка. Они представляют собой вкусовые «почки». Чувствительные клетки в них окружены опорными и погружены в глубину. Небольшие углубления над ними заполнены слизью, в которую выстоят чувствительные волоски. Они воспринимают раздражение от веществ, имеющих к ним стереохимическое сродство. Нервные волокна, отходящие от почек, формируют вкусовые нервы — веточки п. glossopharyngeus, п. Hngualis, chorda tyrapani. Импульсы поступают в ядра одиночного пучка продолговатого мозга, отсюда нейроны передают импульсы в составе медиальной петли в ядра таламуса. Нейроны, заложенные здесь, передают импульсы в кору. Различают вкусовые ощущения следующих типов: сладкий, кислый, соленый, горький. Всевозможные оттенки вкусовых ощущений зависят от множества дополнительных вкусовых и обонятельных раздражений, создаваемых определенными веществами. Обонятельный и вкусовой анализаторы тесно связаны в своей активности. Оба они принадлежат к легкоадаптирующим- ся. Кроме того, оба могут поддаваться «тренировке» — понижению порогов возбуждения и повышению чувствительности к определенным факторам.
♦ ***
На основании изложенного резюмируем основные механизмы и принципы, позволяющие мозгу анализировать поступающую информацию:
1. Дивергенция и конвергенция сигналов. Дивергенция приводит к «размыванию» ин формации, к снижению точности восприятия, но позволяет выявить сигнал. Конвергенция позволяет сузить поток информации.
2. Принцип картирования — проецирование в соответственную точку мозга (точка в точку) — например, соматотопическая организация, ретинотопическая организация.
3. Принцип специализации нейрона, вычленяющего отдельные признаки стимула или совокупность этих признаков.
4. Принцип сохранения модальности нейрона. Это один из самых важных принципов, благодаря которому мы можем ощущать кислое и соленое, запах розы и запах гнилостного, различать красное и черное, мелодию и какофонию и т. п.
5. Принцип колоночной обработки информации — в колонке при наличии нейронов раз ной специализации происходит тщательная (какая только возможна у данного человека) обработка информации — причем обязательно с соблюдением принципа — от простого к сложному. Вероятно, у некоторых людей развитие колоночного анализа достигает макси мума. К примеру — художники, способные дифференцировать миллион цветовых отген ков, дегустаторы духов, вин, блюд и т. п.
6. Ассоциативный способ обработки информации: при его отсутствии возникают иллю зии — зрительные, слуховые, тактильные, так как каждый анализатор работает независимо друг от друга. Но взаимная работа, наличие нейронов, анализирующих поступающую ин формацию с «позиций» полимодальности, дает возможность мозгу реально отражать внеш ний мир, несмотря на несовершенство органов чувств. В этом плане важна роль нейронов коры, которые «дорисовывают» события, позволяют видоизменять ощущения в соответст вии с информацией, поступающей от других рецепторов. Поэтому перевернутое изображе ние на сетчатке нейронами коры переворачивается в сознании и возникает правильное пред ставление об окружающем мире. Таково значение взаимодействия различных анализато ров, которое совершается в ассоциативных участках коры с участием структур, отвечаю щих за процессы обобщения (2-я сигнальная система действительности по Павлову).
Замечание по процессам адаптации анализатора
Существуют центральные и периферические механизмы адаптации, т. е. утраты чувствительности анализатора. О периферических механизмах уже говорилось выше. Например, рецепторы прикосновения и рецепторы вибрации — это типичный пример быстроадапти- рующихся рецепторов. Но есть центральные механизмы, позволяющие регулировать чувствительность анализаторов. Например, адаптация к запахам обусловлена не развитием адаптации в рецепторах, а развитием торможения в центральных нервных сетях обонятельного пути. Обусловлено это тем, что в обонятельных луковицах, где идет обработка информации от обонятельных рецепторов, имеются тормозные клетки, активация которых со стороны вышележащих структур мозга приводит к блокаде проведения импульсов от обонятельных рецепторов. В слуховом анализаторе имеются тормозные волокна, снижающие чувствительность волосковых клеток к звуковой волне.








