 2020-06-29
2020-06-29 248
248Этот вид таутомерии характерен для азотсодержащих гетероциклов с фрагментом N=C—ОН.
Взаимопревращение таутомерных форм связано с переносом протона от гидроксильной группы, напоминающей фенольную ОН-группу, к основному центру — пиридиновому атому азота и наоборот. Обычно лактамная форма в равновесии преобладает.
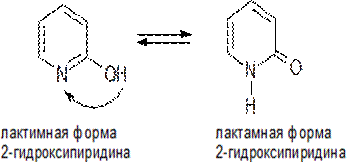
11)Нуклеозиды: строение, номенклатура, гидролиз, характер связей
Нуклеиновые основания образуют связь за счет одного из атомов азота с аномерным центром пентозы (D-рибозы или 2-дезокси-D-рибозы). Этот тип связи аналогичен обычной гликозидной связи и известен как N-гликозидная связь, а сами гликозиды - как N-гликозиды. В химии нуклеиновых кислот их называют нуклеозидами.
В состав природных нуклеозидов пентозы входят в фуранозной форме (атомы углерода в них нумеруют цифрой со штрихом). Гликозидная связь осуществляется с атомом азота N-1 пиримидинового и N-9 пуринового оснований.
Природные нуклеозиды всегда являются β-аномерами.
В зависимости от природы углеводного остатка различают рибонуклеозиды и дезоксирибонуклеозиды. Для нуклеозидов употребительны названия, производимые от тривиального названия соответствующего нуклеинового основания с суффиксами -идин у пиримидиновых и -озин у пуриновых нуклеозидов.
12) Нуклеотиды: строение, номенклатура, характер связи. Гидролиз нуклеотидов.
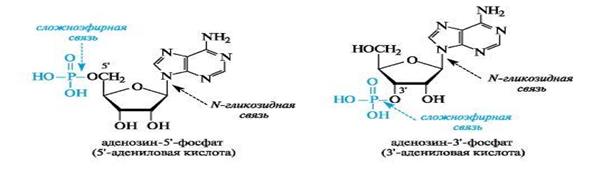
Нуклеотидами называют фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил при С-5' или С-3' в остатке рибозы (рибонуклеотиды) или дезоксирибозы (дезоксирибонуклеотиды).
Общий принцип строения нуклеотидов показан на примере фосфатов аденозина. Для связывания трех компонентов в молекуле нуклеотида используются сложноэфирная и N-гликозидная связи.
Нуклеотиды можно рассматривать, с одной стороны, как эфиры нуклеозидов (фосфаты), а с другой - как кислоты (в связи с наличием остатка фосфорной кислоты).
За счет фосфатного остатка нуклеотиды проявляют свойства двухосновной кислоты и в физиологических условиях при рН ~7 находятся в полностью ионизированном состоянии.
Для нуклеотидов используют два вида названий. Одно включает наименование нуклеозида с указанием положения в нем фосфатного остатка, например, аденозин-3'-фосфат, уридин-5'-фосфат; другое строится с добавлением сочетания -иловая кислота к названию остатка пиримидинового основания, например, 5'-уридиловая кислота, или пуринового основания, например 3'-адениловая кислота.
Используя принятый для нуклеозидов однобуквенный код, 5'-фосфаты записывают с добавлением латинской буквы «р» перед символом нуклеозида, 3'-фосфаты - после символа нуклеозида. Аденозин-5'-фосфат обозначается рА, аденозин-3'-фосфат - Ар и т. п. Эти сокращенные обозначения используют для записи последовательности нуклеотидных остатков в нуклеиновых кислотах. По отношению к свободным нуклеотидам в биохимической литературе широко используют их названия, как монофосфатов с отражением этого признака в сокращенном коде, например АМР (или АМФ) для аденозин-5'-фосфата Гидролиз 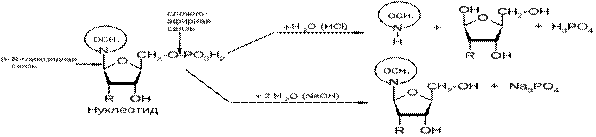
13) Свободные нуклеотиды: цамф и цгмф, атф, адф, фад, над. Строение, функции.
Циклический аденозинмонофосфат (цамф) — производное АТФ, выполняющее в организме роль вторичного посредника, использующегося для внутриклеточного распространения сигналов некоторых гормонов (например, глюкагона или адреналина), которые не могут проходить через клеточную мембрану. Превращает ряд инертных белков в ферменты (цамф-зависимые протеинкиназы), под действием которых происходит ряд биохим. реакций (проведение нервного импульса).
Образование цАМФ стимулируется адреналином.
Циклический гуанозинмонофосфат (цГМФ) - это циклическая форма нуклеотида, образующаяся из гуанозинтрифосфата (GTP) ферментом гуанилатциклазой. Образование стимулируется ацетилхолином.
● цГМФ вовлечен в регуляцию биохимических процессов в живых клетках в качестве вторичного посредника (вторичного мессенджера). Характерно, что многие эффекты цГМФ прямо противоположны цАМФ.
● цГМФ активирует G-киназу и фосфодиэстеразу, гидролизующую цАМФ.
● цГМФ принимает участизе в регуляции клеточного цикла. От соотношения цАМФ/цГМФ зависит выбор клетки: прекратить деление (остановиться в G0 фазе) или продолжить, перейдя в фазу G1.
● цГМФ стимулирует пролиферацию клеток (деление), а цАМФ подавляет
Аденозинтрифосфат (АТФ) - нуклеотид, образованный азотистым основанием аденином, пятиуглеродным сахаром рибозой и тремя остатками фосфорной кислоты. Фосфатные группы в молекуле АТФ соединены между собой высокоэнергетическими (макроэргическими) связями. Связи между фосфатными группами не очень прочные, и при их разрыве выделяется большое количество энергии. В результате гидролитического отщепления от АТФ фосфатной группы образуется аденозиндифосфорная кислота (АДФ) и высвобождается порция энергии.
● Вместе с другими нуклеозидтрифосфатами АТФ является исходным продуктом при синтезе нуклеиновых кислот.
● АТФ отводится важное место в регуляции множества биохимических процессов. Являясь аллостерическим эффектором ряда ферментов, АТФ, присоединяясь к их регуляторным центрам, усиливает или подавляет их активность.
● АТФ является также непосредственным предшественником синтеза циклического аденозинмонофосфата — вторичного посредника передачи в клетку гормонального сигнала.
● Также известна роль АТФ в качестве медиатора в синапсах и сигнального вещества в других межклеточных взаимодействиях
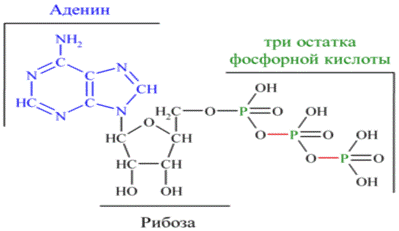
Аденозиндифосфат (АДФ) — нуклеотид, состоящий из аденина, рибозы и двух остатков фосфорной кислоты. АДФ участвует в энергетическом обмене во всех живых организмах, из него образуется АТФ путём фосфорилирования:
АДФ + H3PO4 + энергия → АТФ + H2O.
Циклическое фосфорилирование АДФ и последующее использование АТФ в качестве источника энергии образуют процесс, составляющий суть энергетического обмена (катаболизма).
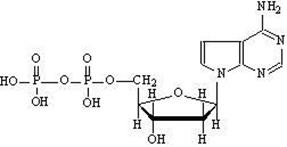
ФАД — флавинадениндинуклеотид — кофермент, принимающий участие во многих окислительно-восстановительных биохимических процессах. ФАД существует в двух формах — окисленной и восстановленной, его биохимическая функция, как правило, заключается в переходе между этими формами.
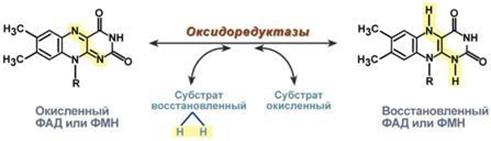
Никотинамидадениндинуклеотид (НАД) - динуклеотид, состоит из двух нуклеотидов, соединённых своими фосфатными группами. Один из нуклеотидов в качестве азотистого основания содержит аденин, другой — никотинамид. Никотинамидадениндинуклеотид существует в двух формах: окисленной (NAD) и восстановленной (NADH).
● В метаболизме NAD задействован в окислительно-восстановительных реакциях, перенося электроны из одной реакции в другую. Таким образом, в клетках NAD находится в двух функциональных состояниях: его окисленная форма, NAD+, является окислителем и забирает электроны от другой молекулы, восстанавливаясь в NADH, который далее служит восстановителем и отдаёт электроны.
● 1. Метаболизм белков, жиров и углеводов. Так как НАД и НАДФ служат коферментами большинства дегидрогеназ, то они участвуют в реакциях
● при синтезе и окислении жирных кислот,
● при синтезе холестерола,
● обмена глутаминовой кислоты и других аминокислот,
● обмена углеводов: пентозофосфатный путь, гликолиз,
● окислительного декарбоксилирования пировиноградной кислоты,
● цикла трикарбоновых кислот.
● 2. НАДН выполняет регулирующую функцию, поскольку является ингибитором некоторых реакций окисления, например, в цикле трикарбоновых кислот.
● 3. Защита наследственной информации – НАД является субстратом поли-АДФ-рибозилирования в процессе сшивки хромосомных разрывов и репарации ДНК, что замедляет некробиоз и апоптоз клеток.
● 4. Защита от свободных радикалов – НАДФН является необходимым компонентом антиоксидантной системы клетки.
14) Первичная структура нуклеиновых кислот. Нуклеотидный состав ДНК и РНК.
Под первичной структурой нуклеиновых кислот понимают порядок, последовательность расположения мононуклеотидов в полинуклеотидной цепи ДНК и РНК. Такая цепь стабилизируется 3',5'-фосфодиэфирными связями.
Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинукпеотидной цепи.
Каждая фосфатная группа в полинукпеотидной цепи, за исключением фосфорного остатка на 5'-конце молекулы, участвует в образовании двух эфирных связей с участием 3'- и 5'-углеродных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3', 5'-фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращённо записывают с помощью однобуквенного кода, например -A-G-C-T-T-A-C-A- от 5'- к 3'-концу.
В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При рН 7 фосфатная группа полностью ионизирована, поэтому нуклеиновые кислоты существуют в виде полианионов (имеют множественный отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной цепи. В РНК, как и в ДНК, нуклеотиды связаны между собой 3',5'-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце - ОН-группа 3'-углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК. Гидроксильная группа у 2'-углеродного атома рибозы делает молекулу РНК нестабильной. Так, в слабощелочной среде молекулы РНК гидролизуются даже при нормальной температуре, тогда как структура цепи ДНК не изменяется.
15) Понятие о вторичной структуре ДНК. Комплементарность нуклеиновых оснований. Водородные связи в комплементарных парах нуклеиновых оснований.
Под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.
Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого -NH...O=C-, а также между амидным и иминным атомами азота -NH...N. Например, как показано ниже, между аденином и тимином образуются две водородные связи, и эти основания составляют комплементарную пару, т. е. аденину в одной цепи будет соответствовать тимин в другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин, между которыми возникают три водородные связи.
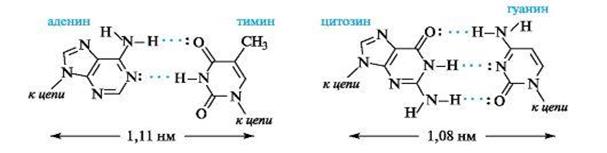
Водородные связи между комплементарными основаниями - один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой. Это означает, что первичная структура, т.е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи.
Комплементарность цепей и последовательность звеньев составляют химическую основу важнейшей функции ДНК - хранения и передачи наследственной информации.
В стабилизации молекулы ДНК наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, их называют стэкинг-взаимодействиями. Таким образом, взаимодействия азотистых оснований между собой скрепляют двойную спираль молекулы ДНК и вдоль, и поперек ее оси.
16) Аминокислоты номенклатура и классификация.
По систематической номенклатуре названия аминокислот образуются из названий соответствующих кислот прибавлением приставки амино и указанием места расположения аминогруппы по отношению к карбоксильной группе.
Организм синтезирует аминокислоты главным образом из пищевых белков. Но есть целая группа аминокислот, которых организм сам синтезировать не может. Эти аминокислоты называют незаменимыми. К ним относятся (валин, лейцин, изолейцин, лизин, треонин, метионин, фенилаланин и триптофан) Такие аминокислоты должны поступать в организм извне.
В настоящее время известно свыше 150 аминокислот, но только 20 из них входят в состав белков.








