 2020-07-12
2020-07-12 921
921а) Сущность понятия «неаллельные гены». Типы взаимодействия между неаллельными генами.
Неаллельные гены – это гены, расположенные в разных участках (локусах) хромосом и контролирующие развитие разных признаков или вариаций одного признака.
Выделяют следующие типы взаимодействия неаллельных генов: комплементарность, эпистаз и полимерию.
б) Комплементарное взаимодействие генов.
Комплементарность – это такой тип взаимодействия неаллельных генов, при котором гены из разных аллельных пар дополняют действие друг друга, приводя к формированию нового признака. Выделяют доминантную или рецессивную комплементарность (т.е. формирование новых признаков связано с взаимодействием доминантных либо только рецессивных аллелей). Например, серая окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) - альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Таким образом, при комплементарном взаимодействии генов расщепление по фенотипу в потомстве при скрещивании двух дигетерозиготных организмов будет составлять 9:7 (доминантная комплементарность), 9:3:3:1 или 9:6:1 (доминантная и рецессивная комплементарность). Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка - интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
в) Эпистатическое взаимодействие генов. Примеры.
Эпистаз – такая форма взаимодействия неаллельных генов между собой, при которой гены одной аллельной пары подавляют действие генов другой аллельной пары. Ген, подавляющий действие другого гена называется эпистатическим геном (супрессором), а подавляемый им ген носит название гипостатического. Выделяют доминантный и рецессивный эпистаз. Доминантный эпистаз – эпистатическое действие оказывает доминантный ген в гомо- или гетерозиготном состоянии (например, окраска оперения у кур). Рецессивный эпистаз – эпистатическое действие оказывает рецессивный ген только в гомозиготном состоянии (например, окраска шерсти у домовой мыши). При доминантном эпистазе при скрещивании двух гомозиготных особей, отличающихся по двум парам неаллельных генов, расщепление по фенотипу среди гибридов второго поколения будет выражаться соотношением 12:3:1 или 13:3. При рецессивном эпистазе это соотношение 9:3:4. Примером доминантного эпистаза может служить наследование окраски оперения у кур.
г) Генетические основы детерминации количественных признаков (полимерия). Примеры.
Полимерия – тип взаимодействия неаллельных генов между собой, при котором несколько неаллельных генов оказывают влияние на формирование одного признака.
Важная особенность полимерии - суммация действия неаллельних генов на развитие количественных признаков. Если при моногенном наследовании признака возможны три варианта "доз" гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до четырех и более. Суммация "доз" полимерных генов обесчивает существования непрерывных рядов количественных изменений. Биологическое значение полимерии заключается еще и в том, что признаки, кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном. Организм без полимерных генов был бы очень неустойчивым: любая мутация или рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет неблагоприятный характер.
Пример: у человека –степень пигментации кожи, определяемая четырьмя парами генов, колеблется от максимально выраженной у гомозигот по доминантным аллелям во всех четырех парах (Р1Р1Р2Р2Р3Р3Р4Р4) до минимальной у гомозигот по рецессивным аллелям (р1р1р2р2р3р3р4р4). При браке двух мулатов, гетерозиготных по всем четырем парам, которые образуют по 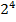 = 16 типов гамет, получается потомство, 1/256 - максимальную пигментацию кожи, 1/256 - минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака.
= 16 типов гамет, получается потомство, 1/256 - максимальную пигментацию кожи, 1/256 - минимальную, а остальные характеризуются промежуточными показателями экспрессивности этого признака.
д) Аддитивный эффект.
Аддитивный эффект – простая суммация действия генов. Предполагается также, что полимерные гены по силе действия равны друг другу. Только при соблюдении этих условий уровень развития количественного признака в потомстве (Fl, F2) будет строго промежуточным между родительскими формами, а кривая распределения генотипов будет точно соответствовать нормальной. Чем больше генов бyдет участвовать в расщеплении, тем меньше в поколении F2 будет доля особей, сходных с исходными родительскими формами, при этом частоты градаций количественного признака будут соответствовать коэффициентам разложения бинома Ньютона.








