2021-10-20
2021-10-20 499
499-избыточное поступление липидов с пищей
-усиление биосинтеза Триглицеридов в печени
-усиление синтеза триглицериидлов в энтероцитах
ОТВЕТ:
1-первое- да -избыточное поступление липидов с пищей, поэтому много всасывается и много синтезируется ХМ, так как они являются транспортной формой экзогенных липидов
2- усиление биосинтеза Триглицеридов в печени нет они же не переносят эндогенные липиды
3- да усиление синтеза триглицериидлов в энтероцитах -оно усилится, если много всасывается, как в первом пункте
4- Хиломикронемия является следствием недостаточности липопротеинлипазы. Накопление в крови хиломикронов. При недостаточной активности липопротеинлипазы крови нарушается переход жирных кислот из хиломикронов (XМ) плазмы крови в жировые депо (не расщепляются триглицериды). лпл же как раз в кровотоке воздействует на ХМ расщепляя их Таги и образуя другие формы ЛП
134 наверное
41. У спортсмена перед ответственным стартом в крови повышается содержание глюкозы до 6,5 мМ/л и уровень СЖК до 1,2 мМ/л (норма 0,4-0,9 мМ/л). В чем причина наблюдаемых изменений?
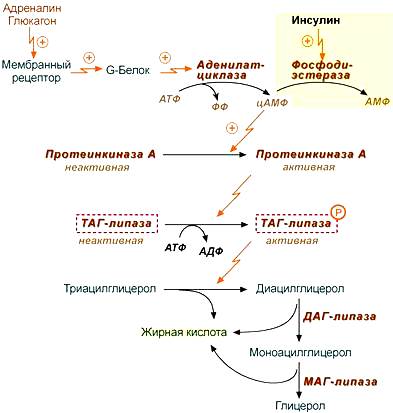
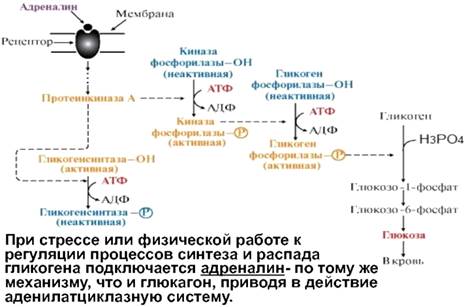
Ответ. Перед стартом повышается уровень адреналина, что приводит к увеличению количества глюкозы и СЖК в крови. Адреналин — катаболический гормон и влияет практически на все виды обмена веществ. Под его влиянием происходит повышение содержания глюкозы в крови и усиление тканевого обмена. Будучи контринсулярным гормоном и воздействуя на β2 адренорецепторы тканей и печени, адреналин усиливает глюконеогенез и гликогенолиз, тормозит синтез гликогена в печени и скелетных мышцах, усиливает захват и утилизацию глюкозы тканями, повышая активность гликолитических ферментов. Также адреналин усиливает липолиз (распад жиров) и тормозит синтез жиров. Это обеспечивается его воздействием на β1 адренорецепторы жировой ткани. В высоких концентрациях адреналин усиливает катаболизм белков.
Активация
-Адреналин связывается с β2-адренорецептором на плазматической мембране клеток печени. В результате связывания лиганда с внешней стороны плазматической мембраны, изменяется конформация всего адренорецептора и активируется сопряженный с адренорецептором, внутриклеточный G-белок.
-В неактивном состоянии G-белок связан с молекулой ГДФ. После активации ГДФ заменяется на ГТФ, а G-белок разделяется на две части.
-Активная часть G-белка присоединяется к ферменту аденилатциклазе и активирует ее. Аденилатциклаза катализирует превращение АТФ в цАМФ.
-цАМФ — является вторичным посредником этой цепи передачи сигнала в клетке. Далее цАМФ распространяется по всей клетке и связывается с цАМФ-зависимой протеинкиназой А, причем с одной молекулой протеинкиназы связвается 4 молекулы цАМФ.
-Активированная протеинкиназа А разделяется на четыре части, две из которых обладают каталитическими активностями. Каждая из каталитических субъединиц способна фосфорилировать киназу-фосфорилазы, активируя ее.
-Наконец, киназа-фосфорилазы фосфорилирует гликоген-фосфорилазу
-Активированная гликоген-фосфорилаза расщепляет гликоген, при этом образуется глюкозо-1-фосфат, которая поступает в кровь.
Особенность этой системы передачи сигнала в клетке состоит в том, что сигнал на большинстве этапов (кроме этапа активации протеинкизаны А молекулами цАМФ) усиливается, например, активированная аденилатциклаза синтезирует множество молекул цАМФ. В результате взаимодействия одной молекулы адреналина с рецептором в плазматической мембране клетки печени, в кровь выводится около 10 миллионов молекул глюкозы