 2014-02-09
2014-02-09 3135
3135Функции сетчатки
В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из белковой части (опсин) и хромофора — 11- цис -ретиналя, под действием фотонов переходящего в транс -ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события: активация родопсина в результате фотоизомеризации ® каталитическая активация G‑белка (Gt, трансдуцин) родопсином ® активация фосфодиэстеразы при связывании с Gta ® гидролиз цГМФ цГМФ–фосфодиэстеразой ® переход цГМФ–зависимых Na+‑каналов из открытого состояния в закрытое ® гиперполяризация плазмолеммы фоторецепторной клетки ® передача сигнала на биполярные клетки.
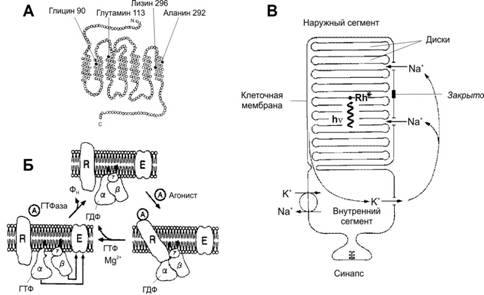
РОДОПСИН И АКТИВАЦИЯ ИОННЫХ КАНАЛОВ [11]. А. Молекула опсина содержит 7 трансмембранных альфа-спиральных участков. Зачернённые кружки соответствуют локализации наиболее распространённых молекулярных дефектов. Так, при одной из мутаций глицин во втором трансмембранном участке в 90-м положении заменён на аспарагин, что приводит к врождённой ночной слепоте. Б. Трансмембранный белок родопсин и его связь с G‑белком (трансдуцин) в плазмолемме фоторецепторной клетки. Возбуждённый фотонами родопсин активирует G‑белок. При этом гуанозиндифосфат, связанный с a‑СЕ G‑белка, заменяется на ГТФ. Отщеплённые a‑СЕ и b‑СЕ действуют на фосфодиэстеразу и заставляют её превращать цГМФ в гуанозинмонофосфат. Это закрывает Na+‑каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; a, b и g — СЕ G‑белка; A — агонист (в данном случае кванты света); E — фермент–эффектор фосфодиэстераза. В. Схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hn) активирует родопсин (Rh*) в дисках, что закрывает Na+‑каналы в клеточной мембране и снижает вход Na+ в клетку.
· Ионные основы фоторецепторных потенциалов
à В темноте Na+‑каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов (рис. 10–5В и 10–6I). Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+,K+–насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca2+ через открытые каналы обеспечивают появление тока (темновой ток).
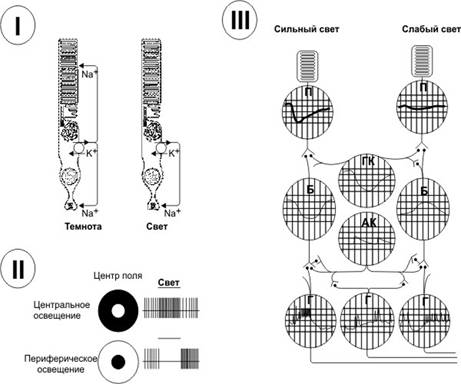
ЭЛЕКТРИЧЕСКИЕ РЕАКЦИИ СЕТЧАТКИ. I. Ответ фоторецептора на освещение. II. Ответы ганглиозных клеток. Освещённые поля показаны белым. III. Локальные потенциалы клеток сетчатки. П — палочки, ГК — горизонтальные клетки, Б — биполярные клетки, AК — амакринные клетки, Г — ганглиозные клетки.
à На свету, т.е. когда свет возбуждает наружный сегмент, Na+‑каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов.
Вышеописанный каскад реакций усиливает световой сигнал и объясняет удивительный факт, каким образом один фотон света — наименьшая возможная квантовая единица световой энергии — может вызывать измеряемый рецепторный потенциал величиной в 1 мВ, а 30 фотонов света могут повышать функциональную готовность палочкового аппарата сетчатки на 50%.
à Возврат к исходному состоянию. Свет, вызывающий каскад реакций, понижающих концентрацию внутриклеточного цГМФ и приводящих к закрытию натриевых каналов, уменьшает содержание в фоторецепторе не только Na+, но и Ca2+. В результате понижения концентрации Ca2+ активируется фермент гуанилатциклаза, синтезирующая цГМФ, и в клетке растёт содержание цГМФ. Это приводит к торможению функций активированной светом фосфодиэстеразы. Оба этих процесса — повышение содержания цГМФ и торможение активности фосфодиэстеразы — возвращают фоторецептор в исходное состояние и открывают Na+‑каналы.
· Световая и темновая адаптация
à Световая адаптация. Если человек длительное время находится в условиях яркого освещения, то в палочках и колбочках происходит превращение значительной части зрительных пигментов в ретиналь и опсин. Большая часть ретиналя превращается в витамин A. Всё это приводит к соответствующему снижению чувствительности глаза, называемому световой адаптацией.
à Темновая адаптация. Напротив, если человек остаётся в темноте продолжительное время, то витамин A вновь превращается в ретиналь, ретиналь и опсин формируют зрительные пигменты. Всё это приводит к повышению чувствительности глаза — темновой адаптации.
Ú При нахождении в темноте световая чувствительность палочек нарастает неравномерно: в первые минуты она увеличивается в десятки раз, в конце первого часа чувствительность палочек к свету возрастает до сотен тысяч раз. В темноте пигменты колбочек восстанавливаются быстрее, чем родопсин палочек, но это не приводит к заметному повышению чувствительности глаза, поскольку абсолютная чувствительность колбочек к свету незначительна.
Ú В дополнение к адаптации, вызываемой изменениями концентрации родопсина и зрительных пигментов, глаз имеет и другие механизмы адаптации. Так, изменение размеров зрачка в течение долей секунды может в 30 раз уменьшить поступление света к сетчатке.
Ú Другой механизм связан с функциональными изменениями нейронов сетчатки. В темноте увеличивается число возбуждённых ганглиозных клеток, что приводит к возрастанию световой чувствительности. ЦНС также влияет на адаптацию сетчатки к действию света. Так, засветка одного глаза понижает чувствительность неосвещённого глаза. Световая чувствительность глаза может изменяться и под воздействием звука.
электрические ответы сетчатки
Различные клетки сетчатки (фоторецепторы, биполярные, горизонтальные, амакринные, а также дендритная зона ганглиозных нейронов) генерируют локальные потенциалы, но не ПД (рис. 10–6). Из всех клеток сетчатки ПД возникают только в аксонах ганглиозных клеток. Суммарные электрические потенциалы сетчатки — электроретинограмма (ЭРГ). ЭРГ регистрируют так: один электрод накладывают на поверхность роговицы, другой — на кожу лица. ЭРГ имеет несколько волн, связанных с возбуждением различных структур сетчатки и суммарно отражает интенсивность и длительность действия света. Данные ЭРГ могут использоваться в диагностических целях при заболеваниях сетчатки
· Связи между клетками сетчатки
à Фоторецепторы образуют синапсы с биполярными нейронами. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном.
à Биполярные нейроны синаптически связаны с ганглиозными и амакринными клетками. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток.
à Горизонтальные клетки формируют синаптические контакты с отростками фоторецепторов. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам. Соседние горизонтальные клетки связаны между собой щелевыми контактами.
à Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками. Эти клетки получают информацию от биполярных нейронов и передают её ганглиозным нейронам. Функции десятков подтипов амакринных клеток полностью не выяснены. Часть клеток участвует в формировании пути палочкового зрения в сетчатке, другие отвечают за начало и окончание световых сигналов, третьи следят за правильностью движения светового пятна. В целом амакринные клетки выполняют функцию вставочных нейронов, осуществляющих первоначальный анализ световых сигналов.
à Ганглиозные клетки — крупные мультиполярные нейроны многих разновидностей. Биполярные и амакринные клетки образуют синапсы на ганглиозных нейронах. Их аксоны образуют зрительный нерв.
à Мюллер овские клетки. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии — мюллеровские клетки. Их ядра расположены на уровне центральной части внутреннего ядерного слоя. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточном пространстве, где концентрация этих ионов при световом раздражении резко увеличивается. Плазматическая мембрана мюллеровских клеток в области ножки характеризуется высокой проницаемостью для ионов K+. Мюллеровская клетка захватывает K+ из наружных слоёв сетчатки и направляет поток этих ионов через свою ножку в жидкость стекловидного тела.
· Нейромедиаторы. Нейроны сетчатки синтезируют ацетилхолин, дофамин, L -глутаминовую кислоту, глицин, g-аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды. Палочки и колбочки в синапсах с биполярными клетками секретируют глутамат. Разные амакринные клетки выделяют ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты. Нейромедиаторы для биполярных и горизонтальных не идентифицированы.
Локальные потенциалы. Ответы палочек, колбочек и горизонтальных клеток являются гиперполяризующими (рис. 10–6,II), ответы биполярных клеток либо гиперполяризующие, либо деполяризующие. Амакринные клетки создают деполяризующие потенциалы.
· Локальные потенциалы колбочек и палочек. Рецепторные потенциалы колбочек и палочек возникают одинаково быстро, но скорость завершения рецепторного потенциала палочек медленнее. Кривая соотношения амплитуды рецепторного потенциала к интенсивности светового стимула имеет одинаковую форму и для палочек, и для колбочек, но палочки во много раз чувствительнее. Ответная реакция палочек пропорциональна интенсивности стимулов на уровне освещения, который ниже порога для колбочек. С другой стороны, колбочки отвечают пропорционально интенсивности стимулов при таком высоком уровне освещения, когда ответы палочек максимальны и не могут больше изменяться. Различия порогов восприятия освещённости колбочками и палочками расширяет диапазон и устойчивость восприятия светового раздражения сетчаткой. Благодаря чрезвычайно низкому порогу восприятия палочки являются детекторами абсолютно малой освещённости, а колбочки реагируют на изменения освещённости в тех пределах, когда палочки уже достигли своего максимума.
· Проведение сигналов от палочек и колбочек. Зрительные пути от палочек и колбочек имеют некоторые отличия. Нейроны и аксоны ганглиозных клеток, проводящие зрительные сигналы от колбочек, значительно толще, чем те, что проводят сигналы от палочек. Скорость проведения сигналов для колбочкового зрения в два раза выше, чем для проведения сигналов палочкового зрения. Для колбочек, эволюционно представляющих молодую и быструю систему проведения, нейронный путь имеет три звена: колбочки ® биполярные клетки ® ганглиозные нейроны. Более древняя палочковая система зрения имеет зрительный путь из 4 звеньев: палочки ® биполярные клетки ® амакринные клетки ® ганглиозные клетки.
Функциональные особенности клеток сетчатки
· Зрительные образы. Сетчатка вовлечена в формирование трёх зрительных образов. Первый образ формируется под действием света на уровне фоторецепторов, превращается во второй образ на уровне биполярных клеток, в ганглиозных нейронах формируется третий образ. В формировании второго образа принимают также участие горизонтальные клетки, а в образовании третьего задействованы амакринные клетки.
· Латеральное торможение — способ усиления зрительного контраста. Латеральное торможение — важнейший элемент деятельности сенсорных систем, позволяющий в сетчатке усиливать явления контраста. В сетчатке латеральное торможение отмечается во всех нейронных слоях, но для горизонтальных клеток оно является их основной функцией. Горизонтальные клетки латерально синаптически связаны с синаптическими участками палочек и колбочек и с дендритами биполярных клеток. В окончаниях горизонтальных клеток выделяется медиатор, который всегда оказывает тормозное влияние. Таким образом, латеральные контакты горизонтальных клеток обеспечивают возникновение латерального торможения и передачу правильного зрительного паттерна в мозг.
à Биполярные клетки реагируют на контрастность изображения. Некоторые биполяры сильнее реагируют на цветной, нежели на чёрно-белый контраст.
à Ганглиозные клетки реагируют на множество свойств зрительного объекта (например, на контрастность изображения, на светлые и тёмные объекты, однородность освещения, цвет объекта, его ориентацию).
· Рецептивные поля. В сетчатке на 100 млн палочек и 3 млн колбочек приходится около 1,6 млн ганглиозных клеток. В среднем на одну ганглиозную клетку конвергирует 60 палочек и 2 колбочки. Существуют большие различия между периферическими и центральными отделами сетчатки в количестве палочек и колбочек, конвергирующих на ганглиозные нейроны. На периферии сетчатки фоторецепторы, связанные с одной ганглиозной клеткой, образуют её рецептивное поле. Перекрытие рецептивных полей различных ганглиозных клеток позволяет повышать световую чувствительность при низком пространственном разрешении. По мере приближения к центральной ямке соотношение палочек и колбочек с ганглиозными клетками становится более упорядоченным, и на каждое нервное волокно приходится всего лишь несколько палочек и колбочек. В области центральной ямки остаются только колбочки (около 35 000), и количество волокон зрительного нерва, выходящих из этой области, равно количеству колбочек. Это создаёт высокую степень остроты зрения по сравнению с относительно слабой остротой зрения на периферии сетчатки. На рис. 10–6,II показаны: слева — диаграммы рецептивных полей, освещённых по центру и по периферии круга, справа — диаграммы частоты ПД, возникающих в аксонах ганглиозных нервных клетках в ответ на освещение. При центральном освещении возбуждённое рецептивное поле вызывает латеральное торможение по периферии: на верхнем рисунке справа частота импульсов в центре намного больше, чем по краям. При засветке рецептивного поля по краям круга импульсация имеется по периферии и отсутствует в центре.
· Ганглиозные клетки разных типов. Ганглиозные клетки в состоянии покоя генерируют спонтанные потенциалы частотой от 5 до 40 Гц, на которые накладываются зрительные сигналы. Известно несколько типов ганглиозных нейронов.
W‑клетки (диаметр перикариона <10 мкм, скорость проведения ПД 8 м/сек) составляют 40% от общего числа всех ганглиозных клеток. W‑клетки имеют обширное рецептивное поле, они получают сигналы от палочек, передаваемые биполярными и амакринными клетками, и ответственны за сумеречное зрение.
X‑клетки (диаметр 10–15 мкм, скорость проведения около 14 м/сек, 55%) имеют небольшое рецептивное поле с дискретной локализацией. Они ответственны за передачу зрительного образа как такового и все виды цветного зрения.
Y‑клетки (диаметр >35 мкм, скорость проведения >50 м/сек, 5%) — самые крупные ганглиозные клетки — имеют обширное дендритное поле и получают сигналы из различных областей сетчатки. Y‑клетки реагируют на быстрые изменения зрительных образов, быстрые движения перед глазами, быстрые изменения интенсивности света. Эти клетки мгновенно сигнализируют в ЦНС, когда в какой-либо части зрительного поля неожиданно появляется новый зрительный образ.
on– и off–ответы. Многие ганглиозные нейроны возбуждаются при изменениях интенсивности освещения. Наблюдается два типа ответов: on–ответ на включение света и off–ответ на выключение света. Эти разные типы ответов появляются соответственно от деполяризованных или гиперполяризованных биполяров.
Цветовоезрение
Характеристики цвета. Цвет имеет три основных показателя: тон (оттенок), интенсивность и насыщение. Для каждого из цветов существует дополнительный (комплементарный) цвет, который, будучи должным образом перемешан с исходным цветом, дает ощущение белого цвета. Чёрный цвет является ощущением, создаваемым отсутствием света. Восприятие белого цвета, любого цвета спектра и даже дополнительных цветов спектра может быть достигнуто смешением в различных пропорциях красного (570 нм), зелёного (535 нм) и голубого (445 нм) цветов. Поэтому красный, зелёный и голубой — первичные (основные) цвета. Восприятие цвета зависит в какой-то мере от цвета других объектов в поле зрения. Например, красный объект кажется красным, если поле освещается зелёным или голубым цветом, и этот же красный объект будет казаться бледно-розовым или белым, если поле будет освещаться красным цветом.
Цветовосприятие — функция колбочек. Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов.
Трихромазия — возможность различать любые цвета — определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц.
Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтанопии и тританопии (от греч. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий]).
Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой Х наследовании генного дефекта.
Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой Х наследование, полиморфизм гена).
Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена.
Полная цветовая слепота — ахромазия — встречается крайне редко. При ахромазии поражается весь колбочковый аппарат, и человек видит все предметы в разных оттенках серого.
Передача цветовых сигналов
Каждая ганглиозная клетка может стимулироваться как отдельными, так и многими колбочками. Когда все три типа колбочек — красные, голубые и зеленые — стимулируют одну и ту же ганглиозную клетку, сигналы, передаваемые через ганглиозную клетку, будут одинаковыми для любого цвета спектра. Эти сигналы не играют роли в определении различных цветов. Все они будут сигналами белого цвета.
Если ганглиозная клетка возбуждается колбочками только одного цвета, то она будет тормозиться возбуждением колбочки другого типа. Это наблюдается для красных и зелёных колбочек. Красные вызывают возбуждение, а зеленые — торможение ганглиозных клеток и наоборот: когда зеленые — возбуждают, то красные — тормозят. Такой же реципрокный тип отношений наблюдается между колбочками голубого, с одной стороны, и комбинацией красных и зелёных колбочек, с другой стороны, вызывая реципрокные (возбуждение–торможение) отношения между голубым и жёлтым цветом.
Механизм антагонистических эффектов следующий: колбочка одного цвета возбуждает ганглиозную клетку через деполяризованную биполярную клетку, а колбочка другого цвета тормозит ту же ганглиозную клетку через гиперполяризованную биполярную клетку.
Нервныепутиицентры
зрительныепути
Зрительные пути подразделяют на старую систему, куда относятся средний мозг и основание переднего мозга, и новую систему (для передачи зрительных сигналов непосредственно в зрительную кору, расположенную в затылочных долях). Новая система фактически отвечает за восприятие всех зрительных образов, цвета и всех форм осознаваемого зрения.
Основной путь к зрительной коре (новая система). Аксоны ганглиозных клеток в составе зрительных нервов и (после перекреста) в составе зрительных трактов достигают латеральных коленчатых тел. При этом волокна от носовой половины сетчатки в зрительном перекресте не переходят на другую сторону. В левом ЛКТ (ипсилатеральном глазу) волокна от носовой половины сетчатки левого глаза и волокна от височной половины сетчатки правого глаза синаптически контактируют с нейронами ЛКТ, аксоны которых образуют коленчато–шпорный тракт (зрительная лучистость). Коленчато-шпорные волокна проходят к первичной зрительной коре той же стороны. Аналогично организованы пути от правого глаза.
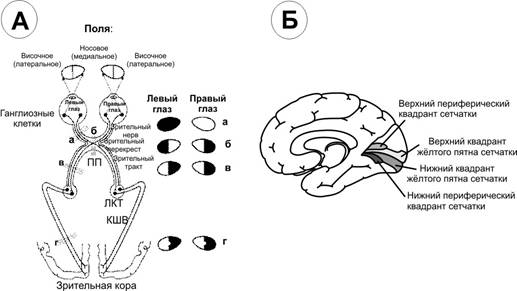
Зрительныепути ( А ) икорковыецентры ( Б ). А. Области перерезки зрительных путей обозначены сточными буквами, а возникающие после перерезки дефекты зрения показаны справа. ПП — перекрест зрительного нерва, ЛКТ — латеральное коленчатое тело, КШВ — коленчато–шпорные волокна Б. Медиальная поверхность правого полушария с проекцией сетчатки в области шпорной борозды.
Другие пути (старая система). Аксоны ганглиозных нейронов сетчатки проходят также в некоторые древние области мозга: к надперекрестным ядрам гипоталамуса (контроль и синхронизация циркадных ритмов); в ядра покрышки (рефлекторные движения глаз при фокусировании объекта, активация зрачкового рефлекса); в верхнее двухолмие (контроль быстрых направленных движений обоих глаз); в ЛКТ и окружающие их области (контроль поведенческих реакций).
Латеральное коленчатое тело (ЛКТ) — часть новой зрительной системы, где оканчиваются все волокна, проходящие в составе зрительного тракта. ЛКТ выполняет функцию передачи информации из зрительного тракта к зрительной коре, в точности сохраняя топологию (пространственное расположение) разного уровня путей из сетчатки. Другая функция ЛКТ заключается в контроле над количеством информации, поступающей к коре. Сигналы для осуществления ЛКТ входного контроля поступают в ЛКТ в виде обратной импульсации из первичной зрительной коры и из ретикулярной области среднего мозга.
зрительнаякора
Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды. Подобно другим частям новой коры, зрительная кора состоит из шести слоев, волокна коленчато–шпорного пути заканчиваются преимущественно на нейронах слоя IV. Этот слой подразделяется на подслои, принимающие волокна от ганглиозных клеток типа Y и X. В первичной зрительной коре (поле 17 по Бродманну) и зрительной области II (поле 18) осуществляется анализ трёхмерного расположения объектов, величины объектов, детализация предметов и их окраски, движения объектов и т.д.
Колонки и полоски. Зрительная кора содержит несколько миллионов вертикальных первичных колонок, каждая колонка имеет диаметр от 30 до 50 мкм и содержит около 1000 нейронов. Нейронные колонки формируют переплетённые между собой полоски шириной в 0,5 мм.
Цветовые колонкообразные структуры. Среди первичных зрительных колонок распределяются вторичные области — колонкоподобные образования («цветовые сгустки»). «Цветовые сгустки» получают сигналы от прилежащих колонок и специфически активируются цветовыми сигналами.
Взаимодействие зрительных сигналов из двух глаз. Зрительные сигналы, поступающие в мозг, остаются раздельными до их вхождения в слой IV первичной зрительной коры. Сигналы из одного глаза входят в колонки каждой полоски, то же самое происходит с сигналами из другого глаза. В ходе взаимодействия зрительных сигналов зрительная кора расшифровывает расположение двух зрительных образов, находит их корреспондирующие точки (точки в одинаковых участках сетчатки обоих глаз) и приспосабливает расшифрованную информацию для определения расстояния до объектов.
Специализация нейронов. В колонках зрительной коры имеются нейроны, выполняющие совершенно определённые функции [например, анализ контраста (в том числе цветового), границ и направлений линий зрительного образа и др.].
В основе детекторной функции нейронов лежит, прежде всего, анализ контраста зрительного образа. Первоначально выявляются границы, степень и градиент контраста, после чего другие нейроны детализируют зрительный образ объекта. Располагающиеся в наружных слоях зрительной коры нейроны являются детекторами линий, определённой длины, углов и других форм зрительных образов. Некоторые группы нейронов определяют границы линий и их направление (детекторы ориентаций). Это простые клетки. Другие клетки определяют направление линий при их вертикальном или боковом смещении (комплексные клетки).
Цвет определяется тем же способом, каким детектируются линии, то есть посредством цветового контраста. Например, нейронная область для красного цвета контрастирует с областью для зелёного цвета, область голубого цвета контрастирует с областью красного и т.д. Все цвета контрастируют против белого цвета.
Механизм цветового контраста зависит от того, что контрастирующие (оппонентные) цвета обоюдно возбуждают специфические нейроны. Полагают, что начальные детали цветового контраста детектируются простыми клетками, в то время как более сложные цветовые контрасты определяются комплексами или даже сверхкомплексами нейронов.
Удаление первичной зрительной коры у человека вызывает потерю восприятия осознаваемых зрительных образов, то есть слепоту. Однако такие слепые люди подсознательно реагируют на изменения интенсивности освещения, передвижения на зрительной сцене и даже некоторые большие зрительные образы. Эти реакции включают повороты глаз, повороты головы, избегание опасных объектов. Такое зрение поддерживается нейронными системами, проходящими из зрительных трактов в верхнее двухолмие и другие отделы старой зрительной системы.
Свойствазрительнойсистемы
Движенияглаз
Наружные мышцы глазного яблока. Движения глаз осуществляют шесть пар поперечно–полосатых мышц, координируемых головным мозгом посредством III, IV, VI пар черепных нервов. Если прямая латеральная мышца одного глаза сокращается, то прямая медиальная мышца другого глаза сокращается на такую же величину. Прямые верхние мышцы работают вместе и перемещают глаза назад, чтобы можно было смотреть вверх. Прямые нижние мышцы дают возможность смотреть вниз. Косая верхняя мышца вращает глаз вниз и наружу, а косая нижняя мышца — вверх и наружу.
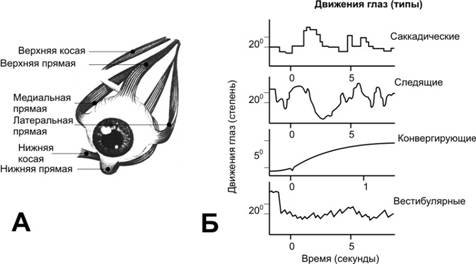
Наружныеглазныемышцы.А. Глазные мышцы левого глаза. Б. Типы движений глаз.
Конвергенция. Одновременное и содружественное движение обоих глаз позволяет, рассматривая близкие предметы, сводить их (конвергенция).
Дивергенция. Рассматривание далёких объектов приводит к разведению зрительных осей обоих глаз (дивергенция).
Диплопия. Поскольку основная часть зрительного поля бинокулярна, ясно, что необходима высокая степень координации движений обоих глаз для удержания зрительного образа на корреспондирующих точках обеих сетчаток и тем самым избежать двоения в глазах (диплопия).
Типы движений. Имеется 4 типа движений глаз (рис. 10–8Б).
Саккады — неощущаемые быстрые скачки (в сотые доли секунды) глаза, прослеживающие контуры изображения. Саккадические движения поддерживают удержание изображения на сетчатке, что достигается периодическим смещением изображения по сетчатке, приводящим к активации новых фоторецепторов и новых ганглиозных клеток.
Плавные следящие движения глаза за движущимся объектом.
Конвергирующие движения — сведение зрительных осей навстречу друг другу при рассматривании объекта вблизи от наблюдателя. Каждый тип движений контролируется нервным аппаратом раздельно, но в конечном итоге все влияния заканчиваются на мотонейронах, иннервирующих наружные мышцы глаза.
Вестибулярные движения глаза — регулирующий механизм, появляющийся при возбуждении рецепторов полукружных каналов и поддерживающий фиксацию взора во время движений головы.
Физиологический нистагм. Даже в условиях, когда субъект пытается фиксировать взглядом неподвижный предмет, глазное яблоко продолжает осуществлять скачкообразные и другие движения (физиологический нистагм). Другими словами, нервно–мышечный аппарат глаза берёт на себя функцию удержания зрительного образа на сетчатке, так как попытка удержать зрительный образ неподвижно на сетчатке приводит к его исчезновению из поля зрения. Именно поэтому необходимость постоянного удержания объекта в поле зрения требует постоянного и быстрого смещения зрительного образа по сетчатке.








