 2015-01-13
2015-01-13 1542
1542Ауксины. Химическая природа, биосинтез, физиологическая роль, практическое применение.
Основным ауксином является индолил-3-уксусная кислота (ИУК). Обнаружены и другие природные вещества с ауксиновой активностью, такие как индолил-3-ацетонитрил, 4-хлор-3-индолилуксусная и фенилуксусная кислоты, однако их ауксиновая активность существенно слабее.
ИУК синтезируется из триптофана в верхушках побега и перемещается сверху вниз по паренхимным клеткам со скоростью 10-15 мм/час благодаря особому механизму полярного транспорта. Возможно также более быстрое передвижение ауксинов по транспортным каналам растения.
Ауксины обладают многообразным физиологическим действием и жизненно важны для роста и развития растений.
Ауксины необходимы для деления и растяжения клеток, для формирования проводящих пучков и корней, способствуют разрастанию околоплодника.
Ауксины обусловливают явление апикального доминирования, т. е. тормозящее действие апикальной почки на рост пазушных почек.
Ауксины играют первостепенную роль в ростовых движениях: фото- и геотропизме и настиях. Ауксины усиливают аттрагирующее действие органов и тканей (т. е. их способность притягивать питательные вещества) и во многих случаях задерживают их старение.
При реализации многих физиологических программ ауксины взаимодействуют с цитокининами и другими фитогормонами.
При высоких концентрациях ауксины повышают образование своего антагониста -- фитогормона этилена.
Первичное действие ауксинов направлено на изменение активности (активацию или репрессию) определенного набора компетентных генов, характерного для данной ткани. ИУК также активирует АТФазу плазмалеммы, вызывая выкачивание протонов из клетки и закисление клеточной стенки. Это приводит к размягчению матрикса стенки, что делает возможным рост клеток растяжением.
На практике ауксины и их синтетические аналоги применяют для размножения клеток и растений в стерильной культуре и для получения трансгенных растений (совместно с цитокининами). Их часто используют для предотвращения предуборочного опадения плодов древесных культур, получения бессемянных плодов томатов, огурцов, баклажанов, перца и др., а также как стимуляторы корнеобразования у черенков; в высоких дозах - как гербициды и дефолианты (2,4-дихлорфеноксиуксусная кислота и др.).
Ауксины синтезируются многими фитопатогенными и симбиотическими микроорганизмами, что помогает последним воздействовать на клетки растения-хозяина.
Аэробная дыхательная цепь. Окислительное фосфолирование.
2. 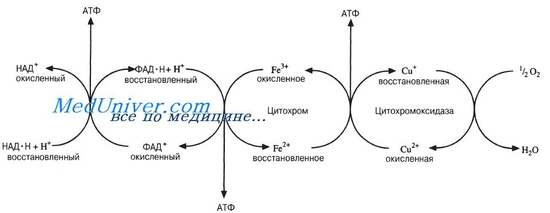
Схема дыхательной цепи.
В процессе окисления пировиноградной кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем рассматривать как 2Н+ + 2е. Именно в таком виде они, акцептированные НАД и ФАД, передаются по цепи переносчиков.
В процессе переноса протонов и электронов важную роль играют ферменты, относящиеся к классу оксидоредуктаз. Оксидоредуктазы, участвующие в дыхательной цепи, делятся на следующие основные группы.
Пиридиновые дегидрогеназы, у которых коферментом служит НАД или НАДФ, отнимают два протона и два электрона от субстрата. При этом к коферментам присоединяются один протон и два электрона. Протон и один электрон связываются с атомом углерода в молекуле НАД, а второй электрон нейтрализует положительный заряд атома азота.
Один протон выделяется в среду. НАД+ и НАДН хорошо растворимы в воде и присутствуют в цитоплазме и митохондриях. Коферменты НАД и НАДФ связаны с ферментом с помощью ионов металла и сульфгидрильных группировок. В зависимости от белкового носителя, к которому присоединен кофермент (НАД или НАДФ), различают более 150 пиридиновых дегидрогеназ. Каждая из них специфична по отношению к определенному субстрату. Необходимо учитывать, что НАД и НАДФ могут воспринимать протоны и электроны лишь в том случае, если субстрат имеет более отрицательное значение потенциала по сравнению с ними.
Окислительное фосфорилирование – процесс переноса неорганического фосфата с макроэргических соединений на АДФ и образование АТФ.








