 2015-02-14
2015-02-14 1577
1577Гипофиз представляет собой эндокринный орган, в котором объединены одновременно три железы, соответствующие его отделам или долям. Передняя доля гипофиза получила название аденогипофиза. По морфологическим критериям это железа эпителиального происхождения, содержащая несколько типов эндокринных клеток. Задняя доля гипофиза, или нейрогипофиз, образуется в эмбриогенезе как выпячивание вентрального гипоталамуса и имеет общее с ним нейроэктодер-мальное происхождение. В нейрогипофизе локализованы веретенообразные клетки — питуициды и аксоны гипоталамических нейронов. Третья, или промежуточная доля гипофиза, как и передняя —эпителиального происхождения, у человека практически отсутствует, но отчетливо выражена, например, у грызунов, мелкого и крупного рогатого скота. У человека функцию промежуточной доли гипофиза выполняет небольшая группа клеток передней части задней доли, эмбриологически и функционально связанных с аденогипофизом.
Кровоснабжение аденогипофиза и нейрогипофиза характеризуется особенностями, во многом определяющими их функции. Артериальные ветви внутренней сонной артерии и вилизиевого круга образуют верхнюю и нижнюю гипофизарные артерии. Верхняя гипофизарная артерия образует мощное капиллярное сплетение в срединном возвышении гипоталамуса, капилляры сливаясь формируют несколько длинных воротных вен, по ножке гипофиза спускающихся в адено-гипофиз и вновь образующих в передней доле сеть синусоидальных капилляров. Следовательно, передняя доля гипофиза прямого артериального снабжения не получает, а кровь в нее поступает из срединного возвышения через воротную систему гипофиза. Эти особенности кровоснабжения гипофиза играю основную роль в регуляции функций передней доли, поскольку аксоны нейросекреторных клеток гипоталамуса в области срединного возвышения образуют
169
 Таблица 5.1. Органы, ткани и клетки с эндокринной функцией
Таблица 5.1. Органы, ткани и клетки с эндокринной функцией
| № | Органы | Ткань, клетки | Гормоны |
| п/п | |||
| 1. ЭНДОКРИННЫЕ ЖЕЛЕЗЬ | |||
| 1. | ГИПОФИЗ | ||
| а) аденогипофиз | Кортикотрофы | Кортикотропин | |
| Меланотропин | |||
| Гонадотрофы | Фоллитропин | ||
| Лютропин | |||
| Тиреотрофы | Тиреотропин | ||
| Соматотрофы | Соматотропин | ||
| Лактотрофы | Пролактин | ||
| б) нейрогипофиз | Питуициты | Вазопрессин | |
| Окситоцин | |||
| 2. | НАДПОЧЕЧНИКИ | ||
| а) корковое вещество | Клубомковая зона | Минералокортикоиды | |
| Пучковая зона | Глюкокортикоиды | ||
| Сетчатая зона | Половые стероиды | ||
| б) мозговое вещество | Хромаффинные | Адреналин | |
| клетки | (Норадреналин) | ||
| 3. | ЩИТОВИДНАЯ ЖЕЛЕЗА | Фолликулярные | Трийодтиронин |
| тиреоциты | Тетрайодтиронин | ||
| К-клетки | Кальцитонин | ||
| 4. | ОКОЛОЩИТОВИДНЫЕ ЖЕЛЕ | ||
| Главные клетки | Паратирин | ||
| К-клетки | Кальцитонин | ||
| 5. | ЭПИФИЗ | Пинеоциты | Мелатонин |
| 2. ОРГАНЫ | С ЭНДОКРИННОЙ ТКАНЬЮ | ||
| 6. | ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА | ||
| Островки Лангерганса | |||
| альфа-клетки | Глюкагон | ||
| бета-клетки | Инсулин | ||
| дельта-клетки | Соматостатин | ||
| 7. | ПОЛОВЫЕ ЖЕЛЕЗЫ | ||
| а) семенники | Клетки Лейдига | Тестостерон | |
| Клетки Сертоли | Эстрогены | ||
| Ингибин | |||
| б) яичники | Клетки гранулезы | Эстрадиол | |
| Эстрон | |||
| Прогестерон | |||
| Желтое тело | Прогестерон | ||
| 3. ОРГАНЫ С ИНКРЕТОРНОЙ ФУНКЦИЕЙ КЛЕТОК | |||
| 8. | ПЛАЦЕНТА | Синцитиотрофобласт | Хорионический |
| гонадотропин | |||
| Пролактин | |||
| Цитотрофобласт | Эстриол | ||
| Прогестерон | |||
| 9. | ТИМУС | Тимоциты | Тимозин, Тимопоэтин |
| 10. | ПОЧКА | ЮГА | Ренин |
| Интерстиций | Эритропоэтин | ||
| Канальцы | Кальцитриол | ||
| 11. | СЕРДЦЕ | Миоциты предсердий | Атриопептид |
| Соматостатин | |||
| Ангиотензин-И |
170

 аксовазальные контакты и нейросекрет с регуляторными пептидами через воротные сосуды поступает в аденогипофиз. Задняя доля гипофиза получает артериальную кровь из нижней гипофизарной артерии. Наиболее интенсивный кровоток имеет место в аденогипо-физе, при этом его уровень (0,8 мл/г/мин) выше, чем в большинстве других тканей организма.
аксовазальные контакты и нейросекрет с регуляторными пептидами через воротные сосуды поступает в аденогипофиз. Задняя доля гипофиза получает артериальную кровь из нижней гипофизарной артерии. Наиболее интенсивный кровоток имеет место в аденогипо-физе, при этом его уровень (0,8 мл/г/мин) выше, чем в большинстве других тканей организма.
Венозные сосуды аденогипофиза впадают в венулы нейрогипофиза. Венозный отток из гипофиза осуществляется в пещеристый венозный синус твердой мозговой оболочки (меньшая часть) и далее в общий кровоток. Большая часть крови оттекает ретроградно в срединное возвышение, что играет определяющую роль в реализации механизмов обратной связи между гипофизом и гипоталамусом. Артериальные сосуды гипофиза получают симпатическую иннервацию по постгангли-онарным волокнам, тянущимся вдоль сосудистой сети.
Функции аденогипофиза. Структура передней доли гипофиза представлена 8 типами клеток, из которых основная секреторная функция присуща хромафильным клеткам 5 групп. Выделяют следующие типы клеток: 1) ацидофильные красные клетки с мелкими гранулами или соматотрофы — вырабатывают соматотропин (СТГ, гормон роста); 2) ацидофильные желтые клетки с крупными гранулами или лактотрофы — вырабатывают пролактин; 3) базофильные тиреотрофы — вырабатывают тиреотропин (тиреотропный гормон — ТТГ); 4) базофильные гонадотрофы —вырабатывают гонадотропины: фоллитропин (фолликулостимулирующий гормон — ФСГ) и лютро-пин (лютеинизирующий гормон — Л Г); 5) базофильные кортикотро-фы —вырабатывают кортикотропин (адренокортикотропный гормон — АКТГ). Кроме того, также как и в клетках промежуточной доли, в базофильных кортикотрофах образуются бета- эндорфин и мелано-тропин, поскольку все эти вещества происходят из общей молекулы предшественника липотропинов.
Таким образом, в аденогипофизе синтезируются и секретируются пять основных типов гормонов: кортикотропин, гонадотропины (фоллитропин и лютропин), тиреотропин, пролактин и соматотропин. Первые три из них обеспечивают гипофизарную регуляцию периферических эндокринных желез (коры надпочечников, половых желез и щитовидной железы), т.е. участвуют в реализации гипофи-зарного пути управления. Для двух других гормонов (соматотропина и пролактина) гипофиз выступает в роли периферической эндокринной железы, поскольку эти гормоны сами действуют на ткани-мишени (рис.5.1.). Регуляция секреции аденогипофизарных гормонов осуществляется с помощью гипоталамических нейропептидов, приносимых кровью воротной системы гипофиза. Регуляторные нейро-пептиды называют "либеринами", если они стимулируют синтез и секрецию аденогипофизарных гормонов, или "статинами", если они останавливают гормональную продукцию аденогипофиза. Не для всех гипофизарных гормонов установлены статины, хотя соматостатин может тормозить продукцию не только соматотропина, но и других гормонов.
171

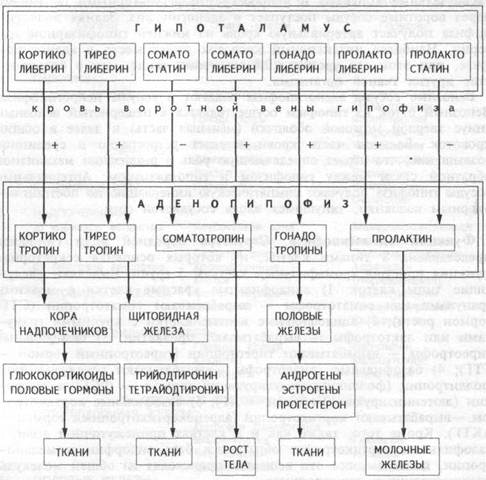
Рис.5.1. Основные гормоны аденогипофиза.
(+) — стимуляция, (-) — подавление секреции.
Регуляция секреции и физиологические эффекты кортикотропина. Кортикотропин является продуктом расщепления крупного (239 аминокислот) гликопротеина проопиомелано-кортина, образующегося базофильными кортикотрофами. Этот белок делится на две части, одна из которых при расщеплении служит источником кортикотропина и меланотропина, а вторая, называемая липотропином, — расщепляясь дает кроме меланотропина морфино-подобный пептид эндорфин, играющий важнейшую роль в антино-цицептиновной (антиболевой) системе мозга и в модуляции секреции гормонов аденогипофиза.
Секреция кортикотропина происходит постоянно пульсирующими вспышками с четкой суточной ритмичностью. Наивысшая концентрация гормона в крови отмечается в утренние часы, а наиболее
172

 низкая — с 22 до 2 часов ночи. Регуляция секреции представлена прямыми и обратными связями. Прямые связи реализуются корти-колиберином гипоталамуса, а обратные запускаются содержанием в крови гипофиза кортикотропина и уровнем гормона коры надпочечников кортизола в системной циркуляции. Обратная связь имеет отрицательную направленность и замыкается как на уровне гипоталамуса (подавление секреции кортиколиберина), так и гипофиза (торможение секреции кортикотропина). Продукция кортикотропина резко возрастает при действии на организм сильных раздражителей, например, холода, боли, физической нагрузки, эмоций, а также под влиянием гипогликемии (снижение сахара в крови).
низкая — с 22 до 2 часов ночи. Регуляция секреции представлена прямыми и обратными связями. Прямые связи реализуются корти-колиберином гипоталамуса, а обратные запускаются содержанием в крови гипофиза кортикотропина и уровнем гормона коры надпочечников кортизола в системной циркуляции. Обратная связь имеет отрицательную направленность и замыкается как на уровне гипоталамуса (подавление секреции кортиколиберина), так и гипофиза (торможение секреции кортикотропина). Продукция кортикотропина резко возрастает при действии на организм сильных раздражителей, например, холода, боли, физической нагрузки, эмоций, а также под влиянием гипогликемии (снижение сахара в крови).
Физиологические эффекты кортикотропина делят на надпочечни-ковые и вненадпочечниковые. Надпочечниковое действие гормона является основным и заключается в стимуляции (через системы аде-нилатциклаза-цАМФ и Са"+) клеток пучковой зоны коры надпочечников, секретирующей глюкокортикоиды (кортизол и кортикосте-рон). Значительно меньший эффект кортикотропин оказывает на клетки клубочковой и пучковой зон коры надпочечников, т.е. на продукцию минералокортикоидов и половых стероидов. Под влиянием кортикотропина усиливается стероидогенез (синтез гормонов), за счет повышения образования и активации транскрипции генов, что при избытке гормона вызывает гипертрофию и гиперплазию коры надпочечников.
Вненадпочечниковое действие кортикотропина заключается в следующих эффектах: 1) липолитическое действие на жировую ткань, 2) повышение секреции инсулина и соматотропина, 3) гипогликемия из- за стимуляции секреции инсулина, 4) повышенное отложение меланина с гиперпигментацией из- за родства молекулы гормона с меланотропином.
Избыток кортикотропина сопровождается развитием гиперкорти-цизма с преимущественным увеличением секреции надпочечниками кортизола и носит название "болезнь Иценко- Кушинга". Основные проявления типичны для избытка глюкокортикоидов. Дефицит кортикотропина ведет к недостаточности глюкокортикоидов с выраженными метаболическими сдвигами и снижением устойчивости организма к влияниям среды.
Регуляция секреции и физиологические эффекты гонадотропинов. Секреция гонадотропинов из специфических гранул гипофизарных клеток имеет четко выраженную цикличность как у мужчин, так и особенно у женщин, о чем речь будет идти в разделе, посвященном половым гормонам. Молекулы гонадотропинов секретируются с прикрепленными на конце углеводных цепей гликопротеида сиаловыми кислотами, что защищает их от разрушения в печени. Как фоллитропин, так и лютропин образуются и секретируются одними и теми же клетками и активация их секреции обеспечивается единым гонадолиберином гипоталамуса. Эффект последнего на секрецию как фоллитропина, так и лютропина, или обоих гонадотропинов вместе, зависит от циклических изменений содержания в крови половых гормонов — эстрогенов, прогестерона
173
 и тестотерона (отрицательная обратная связь). Главный тормозной эффект на продукцию фоллитропина оказывает по механизму обратной связи гормон семенников — ингибин. Тормозит секрецию гонадотропинов гормон аденогипофиза пролактин; выделение лют-ропина угнетают и глюкокортикоиды.
и тестотерона (отрицательная обратная связь). Главный тормозной эффект на продукцию фоллитропина оказывает по механизму обратной связи гормон семенников — ингибин. Тормозит секрецию гонадотропинов гормон аденогипофиза пролактин; выделение лют-ропина угнетают и глюкокортикоиды.
Реализация эффектов гонадотропинов осуществляется через систему аденилатциклаза-цАМФ. Основное действие они оказывают на половые железы, причем не только на образование и секрецию половых гормонов, но и на функции яичников и семенников. Фоллитропин связывается с рецепторами клеток примордиального фолликула в яичниках и клеток Сертоли в семенниках, приводя к четкому морфогенетическому эффекту в виде роста фолликулов яичника и пролиферации клеток гранулезы у женшин, росту яичек, пролиферацию клеток Сертоли и сперматогенезу у мужчин. В продукции половых гормонов фоллитропин оказывает вспомогательный эфо.ект, готовя секреторные структуры к действию лютропина и стимулируя ферменты биосинтеза половых стероидов. Лютропин вызывает овуляцию и рост желтого тела в яичниках, стимулирует клетки Лейдига в семенниках. Он является ключевым гормоном стимуляции образования и секреции половых гормонов: эстрогенов и прогестерона в яичниках, андрогенов в семенниках. Для оптимального развития гонад и секреции половых гормонов необходимо синергичное действие фоллитропина и лютропина, поэтому их часто объединяют единым названием гонадотропины.
Регуляция секреции и физиологические эффекты тиреотропина. Тиреотропин — гликопротеидный гормон аденогипофиза секретируется непрерывно, с четкими колебаниями в течение суток, при этом максимум содержания в крови приходится на часы, предшествующие сну. Секреция тиотропина стимулируется ти-реолиберином гипоталамуса, а подавляется соматостатином. По механизму отрицательной обратной связи регуляция осуществляется содержанием в крови гормонов щитовидной железы (трийодтиронина и тетрайодтиронина), секрецию которых тиреотропин усиливает. Замыкание обратной связи возможно как на уровне гипоталамуса (подавление ^продукции тиреолиберина), так и гипофиза (подавление секреции тиреотропина). Тормозят секрецию тиреотропина и глюкокортикоиды. Тиреотропин секретируется в повышенных количествах при действии на организм низкой температуры, другие же воздействия — травма, боль, наркоз — секрецию гормона подавляют.
Тиреотропин связывается со специфическим рецептором фолликулярных клеток щитовидной железы и вызывает метаболические реакции с помощью четырех вторичных посредников: цАМФ, инози-тол-3-фосфата, диацилглицерола и комплекса Са + кальмодулин. Под влиянием тиреотропина в клетках фолликулов щитовидной железы меняются все виды обмена веществ, ускоряется захват иода и осуществляется синтез тиреоглобулина и тиреоидных гормонов. Тиреотропин увеличивает секрецию гормонов щитовидной железы активацией гидролиза тиреоглобулина. Благодаря увеличению синтеза РНК и белка тиреотропин вызывает увеличение массы щитовидной
174



 железы. Внетиреоидное действие тиреотропина проявляется повышением образования гликозаминогликанов в коже, подкожной и заор-битальной клетчате. Это обычно бывает из-за реализации обратной связи при недостаточной продукции гормонов щитовидной железы, например, при дефиците иода. Избыточная секреция тиреотропина приводит к появлению зоба, гиперфункции щитовидной железы с эффектами избытка тиреоидных гормонов (тиреотоксикоз), пучеглазию (экзофтальм), что в совокупности называют "Базедова болезнь".
железы. Внетиреоидное действие тиреотропина проявляется повышением образования гликозаминогликанов в коже, подкожной и заор-битальной клетчате. Это обычно бывает из-за реализации обратной связи при недостаточной продукции гормонов щитовидной железы, например, при дефиците иода. Избыточная секреция тиреотропина приводит к появлению зоба, гиперфункции щитовидной железы с эффектами избытка тиреоидных гормонов (тиреотоксикоз), пучеглазию (экзофтальм), что в совокупности называют "Базедова болезнь".
Регуляция секреции и физиологические эффекты соматотропина. Соматотропин секретируется аденогипофизар-ными клетками непрерывно и "вспышками" через 20-30 минут с отчетливой суточной ритмикой. Секреция регулируется гипоталами-ческими нейропептидами соматолиберином и соматостатином. Повышение секреции соматотропина происходит во время глубокого сна, на ранних его стадиях (народная мудрость гласит: "человек растет, когда спит"), после мышечных нагрузок, под влиянием травм и инфекций. Стимулируют продукцию соматотропина вазопрессин и эндорфин, а также изменения обмена веществ. Так, гипогликемия активирует секрецию соматолиберина и соматотропина, а гипергликемия — тормозит; избыток аминокислот и снижение свободных жирных кислот в крови активируют секрецию. Эти влияния реализуются через специальные рецепторные нейроны гипоталамуса, воспринимающие сдвиги химизма крови и участвующие в регуляции обмена веществ.
Физиологические эффекты соматотропина связаны с его влияниями на обмен веществ, большинство из которых опосредуется специальными гуморальный факторами (гормонами) печени и костной ткани, получившими название соматомедины (от слова медиатор — посредник). Поскольку эффекты соматомединов на обмен веществ во многом сходны с эффектами инсулина, их нередко еще называют инсулино-подобные факторы роста. Эти эффекты проявляются, в частности, в облегчении утилизации глюкозы тканями, активации в них синтеза белка и жира. Соматомедины опосредуют эффекты соматотропина благодаря специфическим влияниям на хрящевую ткань: стимуляции включения сульфата в синтезируемые протеогли-каны, стимуляции включения тимидина в образуемую ДНК, активации синтеза РНК и белка. В то же время дифференцировка пре-хондроцитов, повышение транспорта аминокислот через их клеточную мембрану обеспечивается не соматомединами, а самим сомато-тропином. Хотя соматомедины и называют инсулиноподобными факторами роста, рецепторы клеточной мембраны для них отличаются от рецепторов инсулина. Описанные эффекты характерны для кратковременного действия соматотропина или ранней фазы его влияния.
При длительной и чрезмерной секреции соматотропина, хотя и сохраняется действие соматомединов на хрящевую ткань, но в целом эффекты соматотропина приобретают четкие контринсулярные черты. Они проявляются в изменениях углеводного и жирового обмена в тканях. Так, соматотропин вызывает гипергликемию из-за
175




 распада гликогена в печени и мышцах и угнетения утилизации глюкозы в тканях, благодаря повышению секреции глкжагона островками Лангерганса поджелудочной железы. Соматотропин увеличивает и секрецию инсулина островками Лангерганса, как за счет прямого стимулирующего действия, так и благодаря гипергликемии. Но в то же время соматотропин активирует инсулиназу печени — фермент, разрушающий инсулин, и вызывает инсулинорезистент-ность тканей. Подобное сочетание стимуляции секреции инсулина с его разрушением и подавлением эффекта в тканях может вести к сахарному диабету, который по происхождению называют гипофи-зарным. Как антогонист инсулина, гипофиз проявляет свои эффекты и в метаболизме липидов. Гормон оказывает пермиссивное (облегчающее) действие по отношению к эффектам катехоламинов и глю-кокортикидов, следствием (его является стимуляция липолиза жировой ткани, повышение уровня свободных жирных кислот в крови, избыточное образование кетоновых тел в печени (кетогенный эффект) и даже жировая инфильтрация печени. Инсулинорезистент-ность тканей может быть связана и с этими сдвигами жирового обмена.
распада гликогена в печени и мышцах и угнетения утилизации глюкозы в тканях, благодаря повышению секреции глкжагона островками Лангерганса поджелудочной железы. Соматотропин увеличивает и секрецию инсулина островками Лангерганса, как за счет прямого стимулирующего действия, так и благодаря гипергликемии. Но в то же время соматотропин активирует инсулиназу печени — фермент, разрушающий инсулин, и вызывает инсулинорезистент-ность тканей. Подобное сочетание стимуляции секреции инсулина с его разрушением и подавлением эффекта в тканях может вести к сахарному диабету, который по происхождению называют гипофи-зарным. Как антогонист инсулина, гипофиз проявляет свои эффекты и в метаболизме липидов. Гормон оказывает пермиссивное (облегчающее) действие по отношению к эффектам катехоламинов и глю-кокортикидов, следствием (его является стимуляция липолиза жировой ткани, повышение уровня свободных жирных кислот в крови, избыточное образование кетоновых тел в печени (кетогенный эффект) и даже жировая инфильтрация печени. Инсулинорезистент-ность тканей может быть связана и с этими сдвигами жирового обмена.
Избыточная секреция соматотропина, если она возникает в раннем детстве, ведет к развитию гигантизма с пропорциональным развитием конечностей и туловища. В юношеском и зрелом возрасте при этом усиливается рост эпифизарных участков костей скелета, зон с незавершенным окостенением, что получило название акромегалия. Растут кисти и стопы, нос, подбородок и т.д. Увеличиваются в размерах и внутренние органы, что называют спланхомега-лия. При врожденном дефиците соматотропина формируется карликовость, называемая "гипофизарный нанизм". После выхода в 1726 г. романа Дж. Свифта "Путешествия Гулливера" таких людей стали называть лилипутами. Приобретенный дефицит гормона в зрелом возрасте выраженного морфогенетического эффекта не вызывает.
Регуляция секреции и физиологические эффекты пролактина. Синтез и секреция аденогипофизом пролактина регулируется гипоталамическими нейропептидами — ингибитором про-лактостатином и стимулятором пролактолиберином. Образование этих гипоталамических пептидов происходит в дофаминергических нейронах гипоталамуса. Секреция пролактина зависит и от уровня в крови эстрогенов, глюкокортикоидов и тиреоидных гормонов.
Основным органом-мишенью пролактина является молочная железа, где гормон стимулирует развитие специфической ткани и лактацию, оказывая свой эффект после связывания со специфическим рецептором с помощью вторичного посредника цАМФ. В молочных железах пролактин влияет именно на процессы образования молока, а не на его выделение. При том гормон стимулирует синтез белка — лактальбумина, а также жиров и углеводов молока. Для регуляции роста и развития молочных желез синергистами пролактина являются эстрогены, но при начавшейся лактаиии эстрогены — антагонисты пролактина. Секреция пролактина стимулируется рефлек-горно актом сосания.

 Кроме влияния на молочные железы пролактин оказывает ряд других эффектов в организме. Он способствует поддержанию секреторной активности желтого тела в яичниках и образованию прогестерона. Пролактин является одним из регуляторов водно-солевого обмена организма, уменьшая экскрецию воды и электролитов, усиливает эффекты альдостерона и вазопрессина, стимулирует рост внутренних органов, эритропоэз, способствует появлению инстинкта материнства. Помимо усиления синтеза белка, пролактин повышает образование жира из углеводов, способствуя послеродовому ожирению.
Кроме влияния на молочные железы пролактин оказывает ряд других эффектов в организме. Он способствует поддержанию секреторной активности желтого тела в яичниках и образованию прогестерона. Пролактин является одним из регуляторов водно-солевого обмена организма, уменьшая экскрецию воды и электролитов, усиливает эффекты альдостерона и вазопрессина, стимулирует рост внутренних органов, эритропоэз, способствует появлению инстинкта материнства. Помимо усиления синтеза белка, пролактин повышает образование жира из углеводов, способствуя послеродовому ожирению.
Функции нейрогипофиза. Нейрогипофиз не образует, а лишь накапливает и секретирует нейрогормоны супраоптического и пара-вентрикулярного ядер гипоталамуса — вазопрессин и окситоцин. Оба гормона находятся в гранулах в связи со специальными белками — нейрофизинами. В процессе секреции содержимое гранул путем эк-зоцитоза поступает в кровь.
Регуляция секреции и физиологические эффекты вазопрессина. Секреция вазопрессина обеспечивается его синтезом в гипоталамических нейронах и регулируется тремя типами стимулов: 1) сдвигами осмотического давления и содержания натрия в крови, воспринимаемыми интероцепторами сосудов и сердца (осмо-, натрио-, волюмо- и механорецепторы), а также непосредственно гипоталамическими нейронами, воспринимающими сдвиг концентрации натрия в крови и микросреде клеток; 2) активацией гипоталамических ядер при эмоциональном и болевом стрессе, физической нагрузке, 3) гормонами плаценты и ангиогензином-II, как содержащимся в крови, так и образуемом в мозге.
Эффекты вазопрессина реализуются за счет связывания пептида б тканях- мишенях с двумя типами рецепторов — V-I и V-2. Стимуляция V-1-реиепторов, локализованных в стенке кровеносных сосудов, через вторичные посредники инозитол- 3- фосфат и кальций-кальмодулин вызывает сужение сосудов, что соответствует названию "вазопрессин". Этот эффект в физиологических условиях выражен слабо из- за низких концентраций гормона в крови. Связывание с V- 2-рецепторами в дисталъных отделах почечных канальцев через вторичный посредник цАМФ вызывает повышение проницаемости стенки канальцев для воды, ее реабсорбцию и концентрирование мочи, что соответствует второму названию вазопрессина —"антидиуретический гормон". Вазопрессин как нейропептид поступает и в ликвор, и по аксонам экстрагипоталамной системы в другие отделы мозга, что обеспечивает его участие в формировании жажды и питьевого поведения, в нейрохимических механизмах памяти.
Недостаток вазопрессина проявляется резко повышенным выделением мочи низкого удельного веса, что называют "несахарным диабетом", а избыток гормона ведет к задержке воды в организме.
Регуляция секреции и физиологические эффекты окситоцина. Синтез окситоцина в гипоталамических нейронах и его секреция нейрогипофизом в кровь стимулируется рефлекторным путем при раздражении рецепторов растяжения матки и механоре-

 цепторов сосков молочных желез. Усиливают секрецию гормона эстрогены. Основные эффекты окситоцина заключаются в стимуляции сокращения матки при родах, сокращении гладких мышц протоков молочных желез, что вызывает выделение молока, а также в регуляции водно- солевого обмена и питьевого поведения. Оскито-цин является одним из дополнительных факторов регуляции секреции гормонов аденогипофиза, наряду с либеринами.
цепторов сосков молочных желез. Усиливают секрецию гормона эстрогены. Основные эффекты окситоцина заключаются в стимуляции сокращения матки при родах, сокращении гладких мышц протоков молочных желез, что вызывает выделение молока, а также в регуляции водно- солевого обмена и питьевого поведения. Оскито-цин является одним из дополнительных факторов регуляции секреции гормонов аденогипофиза, наряду с либеринами.
Гормоны промежуточной доли. Меланотропин у взрослого человека, в отличие от животных с обильным волосяным покровом, практически не синтезируется. Функции этого гормона, заключающиеся в синтезе меланина, его дисперсии в отростках меланоцитов кожи, увеличении свободного пигмента в эпидермисе и в конечном счете повышении пигментации кожи и волос, выполняют в большей мере кортикотропин и липотропин. Выше уже указывалось, что эти гормоны, как и меланотропин, образуются из единого предшественника, в связи с чем при избыточной секреции кортикотропина усиливается пигментация кожи. Усиливают пигментацию кожи и близкие этому гормону пептиды плаценты. Меланотропин тем не менее играет роль как мозговой пептид в нейро-химических процессах памяти (его обнаруживают в аркуатном ядре мозга).
5.2. Функции надпочечников.
Надпочечники являются парной железой внутренней секреции, морфологически и функционально состоящей из двух разных по эмбриональному происхождению тканей — коркового и мозгового вещества Кора надпочечников — производное мезодермы, синтезирует три вида гормонов стероидной природы или кортикостероидов. Мозговое вещество называют также хромаффинной тканью, которая имеет общее с нервной системой происхождение и развивается из эмбриональных симпатических клеток. Кроме надпочечников хромаффинная ткань, образующая катехоламины, обнаруживается в составе симпатического пограничного ствола, брюшном и солнечном сплетении.
Кровоснабжение надпочечника осуществляется тремя группами артерий, отходящих от диафрагмальной и почечной артерий и аорты, а также нескольких добавочных артерий. Ветви этих сосудов образуют раздельные корковые и мозговые артерии, дающие обильные капиллярные сети. Венозный отток крови осуществляется по многочисленным венам в систему нижней полой вены и бассейн воротной вены печени. Как в корковом, так и в мозговом веществе имеется большое количество нервных волокон. Особое значение имеют ветви чревного нерва, несущие пре- и постганглионарные волокна к хромаффинными клеткам мозгового вещества и обеспечивающие поступление к ним регуляторных стимулов.
Функции коры надпочечников. Кора надпочечников, занимающая по объему 80% всей железы, состоит их трех клеточных зон: наружной клубочковой зоны, образующей минералокортикоиды, сред-
178


 ней пучковой зоны, образующей глкжокортикоиды, и внутренней сетчатой зоны, в небольшом количестве продуцирующей половые стероиды. Все кортикоиды образуются из холестерина крови и синтезируемого в самих корковых клетках. При синтезе кортикостеро-идов образуется порядка 50 различных соединений, однако, секре-тируются в кровь в физиологических условиях лишь 7- 8 их них.
ней пучковой зоны, образующей глкжокортикоиды, и внутренней сетчатой зоны, в небольшом количестве продуцирующей половые стероиды. Все кортикоиды образуются из холестерина крови и синтезируемого в самих корковых клетках. При синтезе кортикостеро-идов образуется порядка 50 различных соединений, однако, секре-тируются в кровь в физиологических условиях лишь 7- 8 их них.
Регуляция секреции и физиологические эффекты минералокортикоидов. У человека единственным минерало-кортикоидом, поступающим в кровь, является альдостерон, тогда как у животных (собаки) секретируется и промежуточный продукт — дезоксикортикостерон. Регуляция синтеза и секреции альдостерона осуществляется премушественно ангиотвнзином-П, что дало основание считать альдостерон частью ренин- ангиотензин-альдостеро-новой системы (РААС), обеспечивающей регуляцию водно-солевого обмена и гемодинамики. Поскольку альдостерон регулирует содержание в крови ионов Na+ и К+, обратная связь в регуляции реализуется прямыми эффектами ионов, особенно К+, на клубочковую зону. В РААС обратные связи включаются при сдвигах содержания Na+ в моче дистальных канальцев, объема и давления крови.
Механизм действия альдостерона, как и всех стероидных гормонов, состоит в прямом влиянии на генетический аппарат ядра клеток со стимуляцией синтеза соответствующих РНК, активации синтеза транспортирующих катионы белков и ферментов, а также повышении проницаемости мембран для аминокислот.
Основные физиологические эффекты альдостерона заключаются в поддержании водно-солевого обмена между внешней и внутренней средой организма. Одним из главных органов-мишеней гормона являются почки, где альдостерон вызывает усиленную реабсорбцию натрия в дистальных канальцах с его задержкой в организме и повышении экскреции калия с мочой (см. также главу 12). Под влиянием альдостерона происходит задержка в организме хлоридов и воды, усиленное выведение Н-ионов и аммония, увеличивается объем цирикулирующей крови, формируется сдвиг кислотно-щелочного состояния в сторону алкалоза. Действуя на клетки сосудов и тканей, гормон способствует транспорту натрия и воды во внутриклеточное пространство.
Минералокортикоиды являются жизненноважными гормонами, так как гибель организма после удаления надпочечников можно предотвратить, вводя гормоны извне. Минералокортикоиды усиливают воспаление и реакции иммунной системы. Избыточная их продукция ведет к задержке в организме натрия и воды, отекам и артериальной гипертензии, потере калия и водородных ионов, к нарушениями возбудимости нервной системы и миокарда. Недостаток альдостерона у человека сопровождается уменьшением объема крови, гипотен-зией, угнетением возбудимости нервной системы.
Регуляция секреции и физиологические эффекты глюкокортикоидов. Клетки пучковой зоны секретируют в кровь у здорового человека два основных глюкокортикоида: кортизол и кортикостерон, причем кортизола примерно в 10 раз больше. Ре-
179

 гуляция секреции глюкокортикоидов осуществляется кортикотропи-ном аденогипофиза. Уровень кортизола в крови по обратной связи угнетает секрецию кортиколиберина в гипоталамусе и кортикотро-пина в гипофизе. Секреция глюкокортикоидов происходит непрерывно с четкой суточной ритмикой, при этом максимальные уровни отмечаются в утренние часы, а минимальные — вечером и ночью. Поступающие в кровь гормоны транспортируются к тканям в свободной и связанной с белком (транскортин) формах.
гуляция секреции глюкокортикоидов осуществляется кортикотропи-ном аденогипофиза. Уровень кортизола в крови по обратной связи угнетает секрецию кортиколиберина в гипоталамусе и кортикотро-пина в гипофизе. Секреция глюкокортикоидов происходит непрерывно с четкой суточной ритмикой, при этом максимальные уровни отмечаются в утренние часы, а минимальные — вечером и ночью. Поступающие в кровь гормоны транспортируются к тканям в свободной и связанной с белком (транскортин) формах.
Глюкокортикоиды прямо или опосредованно регулируют почти все виды обмена веществ и физиологические функции. В общем виде метаболические сдвиги можно свести к распаду белков и; липидов в тканях, после чего метаболиты поступают в печень, где из них синтезируется глюкоза, использующаяся как источник энергии. На белковый обмен гормоны оказывают катаболический и антианаболический эффекты, приводя к отрицательному азотистому балансу. Распад белка происходит в мышечной, соединительной и костной тканях, падает уровень альбумина в крови, снижается проницаемость клеточных мембран для аминокислот. Сами гормоны стимулируют катаболизм триглицеридов и подавляют синтез жира из углеводов. Однако из-за гипергликемии и повышения секреции инсулина синтез жира повышается и он откладывается в верхней части туловища, шее и на лице. Эффекты на углеводный обмен в целом противоположны инсулину, поэтому глюкокортикоиды называют контринсулярными гормонами, Гипергликемия под влиянием гормонов возникает за счет усиленного образования глюкозы в печени из аминокислот — глюконеогенеза и подавления утилизации ее тканями. Чувствительность тканей к инсулину глюкокортикоиды снижают, а контринсулярные метаболические эффекты могут вести к развитию стероидного сахарного диабета.
Влияние глюкокортикоидов на реактивность тканей проявляется не только в виде подавления чувствительности к инсулину, но и в повышении чувствительности адренорецепторов к катехоламинам. Глюкокортикоиды вызывают снижение в крови количества лимфоцитов, эозинофилов и базофилов, повышение сенсорной чувствительности и возбудимости нервной системы, поддержание оптимальной регуляции сердечно-сосудистой системы, и, подобно минерало-кортикоидам, задержку натрия и воды при потере калия. Гормоны участвуют в формировании стресса, повышая устойчивость организма к действию чрезмерных раздражителей, подавляют сосудистую проницаемость и воспаление (поэтому их называют адаптивными и противовоспалительными), из-за катаболизма белка в лимфоидной ткани и угнетения иммунных реакций они оказывают антиаллергические эффекты.
Избыток глюкокортикоидов, не связанный с описанной выше усиленной секрецией кортикотропина, получил название синдрома Иценко-Кушинга. Его основные проявления близки болезни Ицен-ко- Кушинга, однако, благодаря обратной связи, избыток глюкокор-тикоидов угнетает секрецию кортикотропина и избыточную пигментацию. Среди проявлений эффектов повышенной секреции гормо-
!80

 нов: мышечная слабость, остеопороз, склонность к сахарному диабету, гипертензия, нарушения половых функций, лимфопения, пеп-тические язвы желудка, изменения психики, предрасположенность к инфекциям. Дефицит глюкокортикоидов вызывает гипогликемию, снижение адренореактивности сердечно- сосудистой системы, замедление сердечного ритма, гигютензию, нейтропению, эозинофилию и лимфоцитоз, снижение сопротивляемости организма инфекциям.
нов: мышечная слабость, остеопороз, склонность к сахарному диабету, гипертензия, нарушения половых функций, лимфопения, пеп-тические язвы желудка, изменения психики, предрасположенность к инфекциям. Дефицит глюкокортикоидов вызывает гипогликемию, снижение адренореактивности сердечно- сосудистой системы, замедление сердечного ритма, гигютензию, нейтропению, эозинофилию и лимфоцитоз, снижение сопротивляемости организма инфекциям.
Регуляция секреции и физиологические эффекты половых стероидов коры надпочечников. Клетками сет чатой зоны у человека секретируются в кровь преимущественно три гормона, относящихся к андрогенам или мужским половым гормонам: андростендион, дегидроэпиандростерон и, существенно меньше, 11-бета-гидроксиандростендион. Наиболее высокий уровень этих гормонов отмечается в 6 часов утра, а наиболее низкий — в 19 часов. Регуляция секреции андрогенов осуществляется с помощью кортикотропина гипофиза.
Физиологические эффекты андрогенов надпочечника проявляются в виде стимуляции окостенения эпифизарных хрящей, повышения синтеза белка (анаболический эффект) в коже, мышечной и костной ткани, а также нормировании у женщин полового поведения. Гормоны являются предшественниками основного андрогена семенников — тестостерона и могут превращаться в него при метаболизме в тканях. Андрогены надпочечников способствуют развитию оволосения по мужскому типу, а их избыток у женщин — к вирилизации, т.е. проявлению мужских черт. Ранний избыток андрогенов ведет к преждевременному проявлению вторичных половых признаков у мальчиков и вирилизации женских половых органов. Нарушения секреции андрогенов коры надпочечников получили название адре-ногенитальных синдромов.
Функции мозгового вещества надпочечников. Мозговое вещество надпочечников содержит хромаффинные клетки, названные так из-за избирательной окраски хромом. По своей сути они являются постган-глионарными нейронами симпатической нервной системы, однако, в отличие от типичный нейронов, клетки надпочечников: 1) синтезируют больше адреналина, а не норадреналина (отношение у человека между ними 6:1); 2) накапливая секрет в гранулах, после поступления нервного стимула они немедленно выбрасывают гормоны в кровь.
Хромаффинные клетки являются частью обшей системы нейро-эндрокринных клеток организма или APUD — системы (Airline and amine Precursors Uptake and Decarboxylation), т.е. системы поглощения и декарбоксилирования аминов и их предшественников. К этой системе относятся нейросекреторные клетки гипоталамуса, клетки желудочно-кишечного тракта (энтериноииты), продуцирующие кишечные гормоны, клетки островков Лангерганса поджелудочной железы и К-клетки щитовидной железы.
Гормоны мозгового вещества — катехоламины — образуются из аминокислоты тирозина поэтапно: тирозин- ДОФА- дофамин- норад-реналин-адреналин. Хотя надпочечник и секретирует значительно
JSI
 больше адреналина, тем не менее в состоянии покоя в крови содержится в четыре раза больше норадреналина, так как он поступает в кровь и из симпатических окончаний. Секреция катехолами-нов в кровь хромаффинными клетками осуществляется с обязательным участием Са+2, кальмодулина и особого белка синексина, обеспечивающего агрегацию отдельных гранул и их связь с фосфо-липидами мембраны клетки.
больше адреналина, тем не менее в состоянии покоя в крови содержится в четыре раза больше норадреналина, так как он поступает в кровь и из симпатических окончаний. Секреция катехолами-нов в кровь хромаффинными клетками осуществляется с обязательным участием Са+2, кальмодулина и особого белка синексина, обеспечивающего агрегацию отдельных гранул и их связь с фосфо-липидами мембраны клетки.
Катехоламины называют гормонами срочного приспособления к действию сверхпороговых раздражителей среды.
Физиологические эффекты катехоламинов, обусловлены различиями в адренорецепторах (альфа и бета) клеточных мембран, при этом адреналин обладает большим сродством к бета-адренорецепторам, а норадреналин — альфа. Основные эффекты стимуляции разных типов адренорецепторов в тканях приведены в табл.5.2.
Таблица 5.2. Основные эффекты стимуляции адренорецепторов катехоламинами


 Альфа-1
Альфа-1
Бета-1
 Сужение артериальных Стимуляция возбудимости,
Сужение артериальных Стимуляция возбудимости,
сосудов проводимости и сократимости
миокарда Сокращение матки
Липолиз в адипозоцитах Расширение зрачка
Гликогенолиз
Расслабление мышц
желудка и кишечника Расслабление мышц желудка
и кишечника
Стимуляция секреции ренина

 Альфа-2
Альфа-2
Бета-2




 Агрегация тромбоцитов Расширение бронхов
Агрегация тромбоцитов Расширение бронхов
Подавление пресинаптического Ррасширение артериальных
освобождения медиатора сосудов
Антилиполиз Расслабление матки
Повышение чувствительности адренорецепторов к адреналину, т.е. пермиссивный эффект, оказывают гормоны щитовидной железы и глюкокортикоиды. Основные функциональные эффекты адреналина проявляются в виде: 1) учащения и усиления сердечных сокращений, 2) сужения сосудов кожи и органов брюшной полости, 3) повышения теплообразования в тканях, 4) ослабления сокращений
182



 желудка и кишечника, 5) расслабления бронхиальной мускулатуры, 6) стимуляции секреции ренина почкой, 7) уменьшения образования мочи, 8) повышения возбудимости нервной системы и эффективности приспособительных реакций. Адреналин вызывает мощные метаболическе эффекты в виде усиленного расщепления гликогена в печени и мышцах из-за активации фосфорилазы, а также подавление синтеза гликогена, угнетение потребления глюкозы тканями, что в целом ведет к гипергликемии. Адреналин вызывает активацию распада жира, мобилизацию в кровь жирных кислот и их окисление. Все эти эффекты противоположны действию инсулина, поэтому адреналин называют контринсулярным гормоном. Адреналин усиливает окислительные процессы в тканях и повышает потребление ими кислорода.
желудка и кишечника, 5) расслабления бронхиальной мускулатуры, 6) стимуляции секреции ренина почкой, 7) уменьшения образования мочи, 8) повышения возбудимости нервной системы и эффективности приспособительных реакций. Адреналин вызывает мощные метаболическе эффекты в виде усиленного расщепления гликогена в печени и мышцах из-за активации фосфорилазы, а также подавление синтеза гликогена, угнетение потребления глюкозы тканями, что в целом ведет к гипергликемии. Адреналин вызывает активацию распада жира, мобилизацию в кровь жирных кислот и их окисление. Все эти эффекты противоположны действию инсулина, поэтому адреналин называют контринсулярным гормоном. Адреналин усиливает окислительные процессы в тканях и повышает потребление ими кислорода.
Таким образом, как кортикостероиды, так и катехоламины обеспечивают активацию приспособительных защитных реакций организма и их энергоснабжение, неспецифически повышая устойчивость к неблагоприятных влияниям среды.

