 2015-03-20
2015-03-20 13101
13101Сон — физиологическое состояние, которое характеризуется прежде всего потерей активных психических связей субъекта с окружающим его миром. Сон является жизненно необходимым для высших животных и человека. Треть жизни человека проходит в состоянии периодически наступающего сна.
Биологическое значение сна. Длительное время считали, что сон представляет собой отдых, необходимый для восстановления энергии клеток мозга после активного бодрствования. Однако, в последнее время биологическое значение сна рассматривается значительно шире. Во-первых, оказалось, что активность мозга во время сна часто выше, чем во время бодрствования. Было установлено, что активность нейронов ряда структур мозга во время сна существенно возрастает. Кроме того, во сне наблюдается активация ряда вегетативных функций. Все это позволило рассматривать сон как активный физиологический процесс, активное состояние жизнедеятельности.
Объективные характеристики (признаки) сна. Сон характеризуется прежде всего потерей активного сознания. Глубоко спящий человек не реагирует на многие воздействия окружающей среды если они не имеют чрезмерной силы. Рефлекторные реакции во время сна снижены. Сон характеризуется фазовыми изменениями ВНД, которые особенно отчетливо проявляются при переходе от бодрствования ко сну.
При переходе от состояния бодрствования ко сну наблюдается следующие фазы:
- уравнительная,
- парадоксальная,
- ультрапарадоксальная,
- наркотическая.
Обычно условно-рефлекторные реакции подчиняются закону силы: на больший по силе условный раздражитель величина условно-рефлекторной реакции больше, чем на слабый раздражитель. Фазы развития сна характеризуются нарушением силовых отношений. Уравнительная фаза характеризуется тем, что животные начинают отвечать одинаковыми по величине условно-рефлекторными ответами на условные раздражители различной силы.
Во время парадоксальной фазы на слабые условные раздражители наблюдается большая по величине условно-рефлекторная реакция, чем на сильные раздражители. Ультрапарадоксальная фаза характеризуется исчезновением условных реакций на положительные условные сигналы и появлением условно-рефлекторного ответа при действии тормозных условных раздражителей. В наркотическую фазу животные перестают отвечать условно-рефлекторной реакцией на любые условные раздражители.
Другим показателем состояния сна является утрата способности к активной целенаправленной деятельности.
Объективные характеристики состояния сна отчетливо обнаруживаются на ЭЭГ и при регистрации ряда вегетативных показателей. Во время сна на ЭЭГ происходит ряд изменений, протекающих в несколько стадий. В состоянии бодрствования характерной является низкоамплитудная высокочастотная ЭЭГ активность (бета-ритм). При закрывании глаз и расслаблении эта активность сменяется альфа-ритмом малой амплитуды. В этот период происходит засыпание человека, он постепенно погружается в бессознательное состояние.
В этот период пробуждение происходит достаточно легко. Через некоторое время альфа-волны начинают складываться в “веретена”. Через 30 минут стадия “веретен” сменяется стадией высокоамплитудных медленных тета-волн. Пробуждение в эту стадию затруднено. Эта стадия сопровождается рядом изменений вегетативных показателей: уменьшается частота сердечных сокращений, снижается кровяное давление, температура тела и др. Стадия тета-волн сменяется стадией высокоамплитудных сверхмедленных дельта-волн. Когда бессознательное состояние становится еще глубже, дельта-волны нарастают по амплитуде и частоте. Дельта-сон — это период глубокого сна. Частота сердечных сокращений, артериальное давление, температура тела в эту фазу достигают минимальных значений.
Описанные изменения ЭЭГ составляют “медленноволновую” стадию сна, она длится 1-1,5 часа. Эта стадия сменяется появлением на ЭЭГ низкоамплитудной высокочастотной активности, характерной для состояния бодрствования (бета-ритм). Так как эта стадия появляется в фазу глубокого сна, она получила название “парадоксального” или “быстроволнового” сна.
Таким образом, по современным представлениям весь период одного цикла сна делится на два состояния, которые сменяют друг друга (такая смена происходит 6-7 раз в течение ночи) и резко отличаются между собой:
- медленноволновый или медленный (ортодоксальный) сон;
- быстрый или парадоксальный сон.
Стадия медленного сна сопровождается высокоамплитудными медленными дельта-волнами в ЭЭГ, а стадия быстрого сна — высокочастотной низкоамплитудной активностью (десинхронизацией), которая характерна для ЭЭГ мозга бодрствующего животного, т. е. по ЭЭГ показателям мозг бодрствует, а организм спит. Это и дало основание назвать эту стадию сна парадоксальным сном.
Если разбудить человека в фазу парадоксального сна, то он сообщает о сновидениях и передает их содержание. Человек, проснувшись в фазу медленного сна, чаще всего не помнит сновидений.
Парадоксальная фаза сна оказалась важной для нормальной жизнедеятельности. Если человека во время сна избирательно лишать только парадоксальной фазы сна, например, будить его как только он переходит в эту фазу, то это приводит к существенным нарушениям психической деятельности. Это свидетельствует о том, что сон и особенно его парадоксальная фаза является необходимым состоянием подготовки к нормальному, активному бодрствованию.
Теории сна.
Гуморальная теория: в качестве причины сна рассматриваются специальные вещества, появляющиеся в крови при бодрствовании. Доказательством этой теории служит эксперимент, При котором бодрствующей собаке переливали кровь животного, лишенного сна в течение суток. Животное — реципиент немедленно засыпало. В настоящее время удалось идентифицировать некоторые гипногенные вещества, например, пептид, вызывающий дельта-сон. Вместе с тем, наличие гипногенных веществ не является фатальным признаком развития сна.
Об этом свидетельствуют наблюдения за поведением двух пар неразделившихся близнецов, У этих близнецов эмбриональное разделение нервной системы произошло полностью, а системы кровообращения имели множество анастомозов. Эти близнецы проявляли различное отношение ко сну: одна девочка, например, могла спать, а другая бодрствовала. Все это указывает на то, что гуморальные факторы не могут рассматриваться как абсолютная причина возникновения сна.
Вторая группа теорий — нервные теории сна. Клинические наблюдения свидетельствовали о том, что при различных опухолевых или инфекционных поражениях подкорковых, особенно стволовых образований мозга, у больных отмечаются различные нарушения сна — от бессонницы до длительного летаргического сна. Эти и другие наблюдения указывали на наличие подкорковых центров сна.
Экспериментально было показано, что при раздражении задних структур субталамуса и гипоталамуса животные немедленно засыпали, а после прекращения раздражения они просыпались. Эти эксперименты указывали на наличие в субталамусе и гипоталамусе центров сна.
В лаборатории И. П. Павлова было установлено, что при применении длительно и настойчиво неподкрепляемого условного раздражителя или при выработке тонкого диффренцировочного условного сигнала, животные, наряду с торможением у них условно-рефлекторной деятельности, засыпали. Эти эксперименты позволили И. П. Павлову рассматривать сон как следствие процессов внутреннего торможения, как углубленное, разлитое, распространившееся на оба полушария и ближайшую подкорку торможение. Так была обоснована корковая теория сна. Однако ряд фактов не могли объяснить ни корковая, ни подкорковая теории сна.
Во-первых, наблюдения за больными, у которых отсутствовали почти все виды чувствительности, показали, что такие больные впадают в состояние сна как только прерывается поток информации от действующих органов чувств. Например, у одного больного из всех органов чувств был сохранен только один глаз, закрытие которого погружало больного в состояние сна. Больная с сохранением чувствительности только на тыльной поверхности предплечья одной руки постоянно пребывала в состоянии сна. Просыпалась она только тогда, когда дотрагивались до участков кожи, сохранивших чувствительность.
Во-вторых, оставалось неясным, почему спят бесполушарные животные и новорожденные дети, у которых кора морфологически еще недостаточно дифференцирована.
Многие вопросы центральной организации процессов сна получили объяснение с открытием восходящих активирующих влияний ретикулярной формации ствола мозга на кору больших полушарий. Экспериментально было доказано, что сон возникает во всех случаях устранения восходящих активирующих влияний ретикулярной формаций на кору мозга.
Наряду с этим были установлены нисходящие влияния коры мозга на подкорковые образования. Особенно важны влияния фронтальных отделов коры больших полушарий на лимбические структуры мозга и гипоталамические центры сна. В бодрствующем состоянии при наличии восходящих активирующих влияний ретикулярной формации на кору мозга, нейроны лобной коры тормозят активность нейронов центра сна заднего гипоталамуса. В состоянии сна, когда снижаются восходящие активирующие влияния ретикулярной формации на кору мозга, тормозные влияния лобной коры на гипоталамические центры сна снижаются.
Важным обстоятельством, имеющим прямое отношение к природе сна, явилось установление факта реципрокных отношений между лимбико-гипоталамическими и ретикулярными структурами мозга. При возбуждении лимбико-гипоталамических структур мозга наблюдается торможение структур ретикулярной формации ствола мозга и, наоборот.
Следовательно, состояния бодрствования и сна характеризуются специфической архитектоникой, своеобразной “раскладкой” корково-подкорковых взаимоотношений.
При бодрствовании за счет потоков афферентации от органов чувств активируются структуры ретикулярной формации ствола мозга, которые оказывают восходящее активирующее влияние на кору больших полушарий. При этом нейроны лобных отделов коры оказывают нисходящие тормозные влияния на центры сна заднего гипоталамуса, что устраняет блокирующие влияния гипоталамических центров сна на ретикулярную формацию среднего мозга.
В состоянии сна при уменьшении потока сенсорной информации снижаются восходящие активирующие влияния ретикулярной формации на кору мозга. В результате чего устраняются тормозные-влияния лобной коры на нейроны центра сна заднего гипоталамуса. Эти нейроны, в свою очередь, начинают еще активнее тормозить ретикулярную формацию ствола мозга. В условиях блокады всех восходящих активирующих влияний подкорковых образований на кору мозга наблюдается медленноволновая стадия сна.
Гипоталамические центры за счет морфофункциональных связей с лимбическими структурами мозга могут оказывать восходящие активирующие влияния на кору мозга при отсутствии влияний ретикулярной формации ствола мозга.
Рассмотренные выше механизмы составляют корково-подкорковую теорию сна, предложенную П. К. Анохиным. Эта теория позволила объяснить все виды сна и его расстройства. Она исходит из ведущего постулата о том, что какова бы ни была причина сна, состояние сна связано с важнейшим механизмом — снижением восходящих активирующих влияний ретикулярной формации на кору мозга.
Развитие сна объясняется уменьшением восходящих активирующих влияний ретикулярной формации за счет торможения активности ее нейронов при электрическом раздражении заднего гипоталамуса.
Сон бескорковых животных и новорожденных детей объясняется слабой выраженностью нисходящих влияний лобной коры на гипоталамические центры сна, которые при этих условиях находятся в активном состоянии и оказывают тормозное действие на нейроны ретикулярной формации Ствола мозга. Сон новорожденного периодически прерывается только возбуждением центра голода, расположенного в латеральных ядрах гипоталамуса, который тормозит активность центра сна. При этом создаются условия для поступления восходящих активирующих влияний ретикулярной формации в кору. Новорожденный просыпается и бодрствует до тех пор, пока не снизится активность центра голода за счет удовлетворения пищевой потребности.
Становится понятным, что во всех случаях резкого ограничения сенсорной информации, имевшего место у некоторых больных, сон возникал вследствие снижения восходящих активирующих влияний ретикулярной формации ствола мозга на кору.
Корково-подкорковая теория сна объясняет многие расстройства сна. Бессонница, например, часто возникает как следствие перевозбуждения коры под влиянием курения, напряженной творческой работы перед сном. При этом усиливаются нисходящие тормозные влияния нейронов лобной коры на гипоталамические центры сна и подавляется механизм их блокирующего действия на ретикулярную формацию ствола мозга.
Неглубокий сон наблюдается при частичной блокаде механизмов восходящих активирующих влияний ретикулярной формации на.кору мозга. Длительный, например, летаргический сон может наблюдаться при раздражении центров сна заднего гипоталамуса сосудистым или опухолевым патологическим процессом. При этом возбужденные клетки центра сна непрерывно оказывают блокирующее влияние на нейроны ретикулярной формации ствола мозга.
Понятие о “сторожевых пунктах” как частичном бодрствовании во время сна объясняется наличием определенных каналов реверберации возбуждений между подкорковыми структурами и корой больших полушарий во время сна на фоне снижения основной массы восходящих активирующих влияний ретикулярной формации на кору мозга. “Сторожевой пункт” или очаг может определяться сигнализацией от внутренних органов, внутренними метаболическими потребностями и внешними жизненно важными обстоятельствами.
Например, кормящая мать может очень крепко спать и не реагировать на достаточно сильные звуки, но она быстро просыпается при легком шевелении новорожденного ребенка. Иногда “сторожевые пункты” могут иметь прогностическое значение. Например, в случае патологических изменений в том или ином органе, усиленная импульсация от него может определять характер сновидений и быть своего рода прогнозом заболевания, субъективные признаки которого еще не воспринимаются в состоянии бодрствования.
Гипнотическое состояние можно определить как частичный сон. Возможно, гипнотическое состояние создается за счет возбуждения лимбико-таламических структур на фоне сохраняющейся части восходящих активирующих влияний ретикулярной формации на кору мозга” определяющих поведенческую деятельность.
Избирательная активация лимбических структур мозга наблюдается при воздействии на мозг импульсов электрического тока при так называемом электросне, при этом формируется гипнозоподобное состояние.
Сон, как особое состояние организма и прежде всего — состояние мозга, характеризуется специфическими корково-подкорковыми соотношениями и продукцией специальных биологически активных веществ, применяется при лечении невротических, астенических состояний, снятия психоэмоционального напряжения и при ряде психосоматических заболеваний (ранние стадии гипертонической болезни, нарушения сердечного ритма, язвенные поражения желудочно-кишечного тракта, кожные и эндокринные расстройства).
Фармакологический сон неадекватен по своим механизмам естественному сну. Различные “снотворные” препараты ограничивают активность разных структур мозга — ретикулярной формации ствола мозга, гипоталамической области, коры головного мозга. При этом нарушаются естественные механизмы формирования стадий сна, его динамики, пробуждения. Помимо этого при фармакологическом сне могут нарушаться процессы консолидации памяти, переработки и усвоения информации и др. Следовательно, использование фармакологических средств для улучшения сна должно иметь достаточное медицинское обоснование.
Современные представления о физиологической архитектуре поведенческого акта (функциональная система поведения). Любая деятельность организма является приспособительной и направлена на достижение организмом полезного приспособительного результата. В основе этой приспособительной деятельности лежит формирование функциональных систем, т. е. совокупности процессов и механизмов, динамически складывающихся для достижения организмом полезного результата. Следовательно, формирование функциональных систем подчинено получению определенного, полезного приспособительного результата. Недостаточный результат может целиком реорганизовать систему, сформировать новую с более совершенным взаимодействием компонентов, обеспечивающих получение полезного результата.
Этапы (узловые механизмы) формирования функциональной системы. Концепция функциональных систем постулирует мысль о том, что среда существования оказывает на организм влияние еще до того, как подействует условный раздражитель. Следовательно, при осуществлении условного рефлекса условный раздражитель действует на фоне так называемой предпусковой интеграции, которая формируется на базе различных видов афферентных возбуждений.
1. Обстановочная афферентация — сумма афферентных возбуждений, возникающих в конкретных условиях существования организма и сигнализирующих об обстановке, в которой пребывает организм.
2. Обстановочная афферентация действует на организм в тот момент, когда в нем имеется тот или иной уровень мотивационного возбуждения (мотивация), находящегося в состоянии скрытого доминирования. Доминирующая мотивация формируется на основе ведущей потребности, при участии мотивационных центров гипоталамуса. Из нескольких потребностей выбирается наиболее актуальная, на базе которой возникает доминирующая мотивация. На стадии афферентного синтеза доминирующая мотивация активирует память.
3. Любая поведенческая реакция, в том числе и условно-рефлекторная возникает быстрее, если подобная ситуация уже встречалась в жизни, т. e. при наличии следов прошлого опыта — памяти. Значение памяти на стадии афферентного синтеза состоит в том, что она извлекает информацию, связанную с удовлетворением доминирующей мотивации.
Эти три вида возбуждений: мотивационное, память и обстановочная афферентация создают предпусковую интеграцию, на фоне которой действует четвертый вид афферентации — пусковая афферентация (пусковой стимул, условный сигнал). Эти четыре вида возбуждений взаимодействуют и обеспечивают формирование Первого этапа, первого узлового механизма функциональной системы поведения — афферентного синтеза (рис. 34).
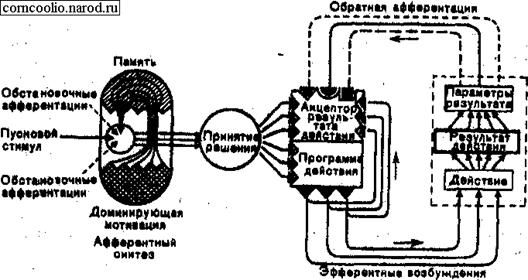
Рис.34. Схема функциональной системы поведенческого акта (по П.К. Анохину).
Основным условием формирования афферентного синтеза является одновременная встреча всех четырех видов афферентаций. Эти виды афферентаций должны обрабатываться одновременно и совместно, что достигается благодаря конвергенции всех видов возбуждений на конвергентных нейронах. Этап афферентного синтеза приводит организм к решению вопроса, какой именно результат должен петь получен в данный момент, он обеспечивает постановку цели, достижению которой будет посвящена вся дальнейшая реализация функциональной системы.
Вторым этапом функциональной системы является принятие решения (постановка цели).
Этот этап характеризуется следующими особенностями:
- Принятие решения осуществляется только на основе полного афферентного синтеза.
- Благодаря принятию решения принимается одна конкретная форма поведения, соответствующая внутренней потребности, прежнему опыту и окружающей обстановке.
- На этапе принятия решения организм освобождается от избыточных степеней свободы, т. е. из сотни возможностей после принятия решения реализуется только одна. Оставшиеся степени свободы дают возможность экономно осуществлять именно то действие, которое должно привести к запрограммированному результату.
- Этап принятия решения способствует формированию интеграла эфферентных возбуждений, в этот период все виды возбуждений приобретают эффекторный, исполнительный характер.
Третьим этапом функциональной системы является формирование программы действия. На данном этапе формируется конкретная цель действия и пути ее реализации. Одновременно с формированием программы действия формируется как бы ее копия, которая сохраняется в нервной системе, в акцепторе результатов действия.
Четвертым этапом формирования функциональной системы является формирование акцептора результатов действия. Это весьма сложный аппарат деятельности мозга, который должен сформировать тонкие нервные механизмы, позволяющие не только прогнозировать признаки (параметры) необходимого в данный момент результата, но и сравнить (сличить) их с параметрами реально полученного результата. Информация о последних приходит к акцептору результатов действия благодаря обратной афферентации. Именно этот аппарат дает возможность организму исправить ошибку поведения или довести несовершенные поведенческие акты до совершенных. Акцептор результатов действия — это идеальный образ будущих результатов действия.
Именно эта модель является эталоном оценки обратных афферентации. Получены данные о том, что в этот нервный комплекс, обладающий высокой степенью мультиконвергентного взаимодействия, приходят возбуждения не только афферентной, но и эфферентной природы. Речь идет о коллатеральных ответвлениях пирамидного тракта, которые через цепь промежуточных нейронов отводят “копии” эфферентных посылок (команд), идущих к эффекторам. Эти эфферентные возбуждения конвергируют на те же промежуточные нейроны сенсомоторной области коры, куда поступают афферентные возбуждения, передающие информацию о параметрах реального результата.
Таким образом, момент принятия решения и начала выхода эфферентных возбуждений из мозга сопровождается формированием обширного комплекса возбуждений, состоящего из афферентных признаков будущего результата и из коллатеральных копий эфферентных возбуждений, поступающих по пирамидному тракту к рабочим аппаратам. К этому же комплексу возбуждений через определенное время присоединяются возбуждения от параметров реально полученного результата. Сам процесс оценки реально полученного результата осуществляется из сличения (сравнения, сопоставления) прогнозированных параметров и параметров реально полученного результата.
Если результаты не соответствуют прогнозу, то в аппарате сличения возникает реакция рассогласования, активирующая ориентировочно-исследовательскую реакцию, которая поднимает ассоциативные возможности мозга на более высокий уровень, тем самым помогает активному подбору дополнительной информации. Именно эта общая активация мозга, реализующаяся в ориентировочно-исследовательской реакции, направляет организм на поиски дополнительной информации. На ее основе формируется более полный афферентный синтез, принимается более адекватное решение, что в свою очередь приводит к формированию более адекватной программы действия и к действию, которое позволяет получить запрограммированный результат.
При достижении желаемого полезного результата в акцепторе результатов действия формируется реакция согласования. В стадию афферентного синтеза поступает санкционирующая афферентация, сигнализирующая об удовлетворении мотивации. На этом функциональная система перестает существовать.
Процессы согласования и рассогласования, возникающие при сличении параметров реально полученного результата с запрограммированным в акцепторе результатов действия, сопровождаются общими реакциями — чувством удовлетворения и неудовлетворения, т.е. положительными и отрицательными эмоциями.
Следовательно, основными этапами, узловыми механизмами функциональной системы являются:
- Афферентный синтез.
- Принятие решения.
- Формирование программы действия.
- Формирование акцептора результатов действия.
- Действие и его результат.
- Сличение параметров результата с их моделью в акцепторе результатов действия, осуществляемое с по мощью обратной афферентации.
Синтез столь разнообразных возбуждений осуществляется на конвергентных нейронах. Именно к ним приходят обстановочная и пусковая афферентации, возбуждение от мотивационных центров. На этих же нейронах осуществляется синтез этих возбуждений со следами ранее протекавших здесь процессов (памятью). Нейроны, на которых формируются механизмы функциональной системы, расположены во всех структурах ЦНС, на всех ее уровнях. Интеграция этих процессов определяет целостную многоуровневую, многокомпонентную приспособительную деятельность организма.








