2015-04-12
2015-04-12 1676
1676Обнародуя свою модель структуры ДНК в 1953 г., Джеймс Уотсон и Фрэнсис Крик писали: «Мы не могли не осознавать, что специфическое спаривание (оснований), постулированное нами, подразумевает наличие какого-то механизма копирования теистического материала». Они же первыми заметили: «Если известен точный порядок оснований в одной из цепей, то можно записать и порядок оснований в другой, поскольку спаривание оснований специфично. Таким образом, одна цепь является комплементом другой; именно это свойство наводит на мысль, что ДНК может удваивать саму себя».
Уотсон и Крик предположили, что для удвоения ДНК должны произойти разрыв водородных связей, удерживающих вместе спиральный дуплекс, и расхождение цепей. Они также высказали мысль, что каждая цепь дуплекса служит матрицейпри синтезе комплементарной цепи, и в результате образуются две пары цепей, в каждой из которых только одна является родительской. Таков механизм точного воспроизведения последовательности нуклеотидных пар в двойной спирали. Уотсон и Крик полагали, что репликация ДНК осуществляется спонтанно, без участия ферментов, но это оказалось неверно. Тем не менее идея о том, что удвоение ДНК происходит путем последовательного соединения нуклеогидов в соответствии с правилом комплементарности, заданным каждой цепью спирали, разрешила концептуальную проблему точного воспроизведения генов.
Согласно общепринятой модели, репликация всех двунитевыных ДНК полуконсервативна. Существуют ли в природе альтернативные способы репликации двунитевой ДНК (например, консервагивный или дисперсный) - неизвестно. Таким образом, после каждого события репликации одна цепь в обеих дочерних молекулах является родительской, консервативной, а другая – новосинтезированной. Именно такой механизм копирования и называется полуконсервативным. Если геном представлен однонитевой ДНК (как в некоторых вирусах), то эта единственная нить служит матрицей для образования комплементарной цепи, с которой она образует дуплекс, а затем на этом дуплексе синтезируются либо дочерние дуплексы, либо однонитевые копии одной из матричных нитей.
Уотсон и Крик уже во второй своей работе 1953 г. предположили возможный механизм копирования наследственного материала. Легко представить, что цепи молекулы ДНК расходятся и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две дочерние двуспиральные молекулы ДНК, не отличимые от родительской, молекулы.
В 1957 г. А. Корнберг обнаружил у бактерии Е. соli фермент, катализирующий процесс полимеризации ДНК из нуклеотидов — ДНК-полимеразу 1. В 1959 г. Артуру Корнбергу (А. Kornberg)) была присуждена Нобелевская премия за открытие механизма биосинтеза ДНК. Он показал, что в основе удвоения молекул ДНК лежат обычные биохимические реакции.
В общем виде реакцию присоединения 5'-дезоксинуклеотидной группы к З'-ОН-группе концевого нуклеотида праймерной цепи можно представить следующим образом:
[dNMP]n + dNTP «[dNMP]n+1 + РРi
где dNMP- любой из четырех обычных нуклеотидов. За один акт репликации нить, содержащая 3’-конец, удлиняется на один остаток, при этом одновременно происходит удаление пирофосфата. Реакция присоединения нуклеотида обратима, но так как неорганический фосфат в клетках быстро разрушается, то реакция активно направлена в сторону синтеза. Репликация ДНК всегда идет от 5’- конца нити ДНК (то есть содержащего 5’-дезоксинуклеотидную группу) к 3’-концу (то содержащему свободную 3-ОН-группу) и нуждается в наличии ранее синтезированного фрагмета нити ДНК в качестве затравки для реакции полимеризации. Такой ДНК-фрагмент, имеющий свободный 3’-конец, называется праймером. Ферменты, катализирующие праймер-зависимую, детерминируемую ДНК-матрицей реакцию присоединения дезоксинуклеотидов, называются ДНК-полимеразами. Несколько различных классов ДНК-полимераз выделены и охарактеризованы, детально описаны свойства этих ферментов и реакции, которые они катализируют. Об их строении и индивидуальных особенностях мы подробно поговорим в следующих главах.
1.2.Вилка репликации. 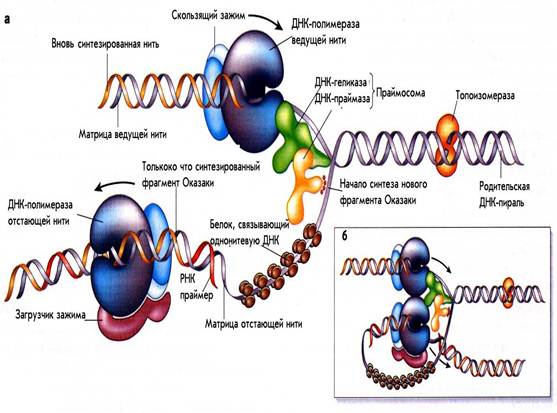
Рис.1. Схема репликативной вилки.
Процесс репликации происхдит в специальных структурах, названных вилками репликации. Схематическое устройство репликативной вилки E.coli представлено на рис. 1. То, что две нити молекулы ДНК расположены антипараллельно друг другу, создает ряд проблем для их олдноаременной разнонаправленной репликации. По мере движения вилки одновременно должны синтезироваться две дочерние цепи. Вилка движется в направлении от 5 ' к 3’ на одной цепи и от 3’ к 5' —- на другой. Однако нуклеиновые кислоты синтезируются только от 5'- к З'-концу. Проблема решается таким образом, что на одной из родительских нитей новая нить синтезируется непрерывно в направлении 5'-3', что совпадает с движением вилки репликации. Это называется лидирующей или ведущей. Другая нить называется отстающей или запаздывающей, так как синтез на ней идет с некорой задержкой по сравнению с лидирующей нитью. Это связано с тем, что ДНК на этой нити синтезируется также от 5' к 3', но в направлении, противоположном движению вилки и короткими фрагментами. Благодаря этому разнонаправленный синтез ДНК может осуществляться в рамках одной структуры – репликативной вилки. Длина таких коротких фрагментов у прокариот составляет 1000-2000 пн. По имени открывшего их ученого они были названы «фрагментами Оказаки». По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются с образованием непрерывной отстающей нити.
Вилка репликации может двигаться как в одну сторону от точки инициации, тогда этот процесс называется однонаправленной репликаций, так и в обе стороны. В таком случае речь идет о двунаправленнгйо репликации. Как это выглядит схематически, показано на рис.2. У эукариот репликация обычно двунапраленная.
Механизмы инициации репликации в точке начала репликации и при образовании фрагментов Оказаки в отстающей цепи в принципе аналогичны, хотя имеются некоторые тонкие различия. В обоих случаях происходит образование коротких РНК-затравок ( праймеров ), комплементарных матричной ДНК, в виде продолжения которых синтезируется новая цепь ДНК. В дальнейшем короткие вставки РНК замещаются сегментами ДНК, которые затем объединяются с образованием непрерывной отстающей нити.
Все живые организмы на Земле обычно делят на прокариот и эукариот (от греч. карион — ядро). Главной особенностью прокариот является отсутствие у них в отличие от эукариот полноценного клеточного ядра, покрытого оболочкой. Генетический материал прокариот расположен в нуклеоиде - примитивном эквиваленте ядра эукариот. Клетки прокариот имеют очень небольшие размеры — около 1 мкм. Объем эукариотических клегок в 800-1000 раз больше объема клеток прокариот. К прокариотам относятся бактерии и археи (или архебактерии), предки которых возникли около 4 млрд лет назад. Эукариоты могут быть как одноклеточными, гак и многоклеточными. Они появились па Земле примерно через 500 млн лет после прокариот.
По современным представлениям ДНК-метаболизм у прокариот имеет некоторые отличия от такового эукариот. Описывая процессы репликации и рекомбинации мы будем каждый раз подчеркивать эти отличия.
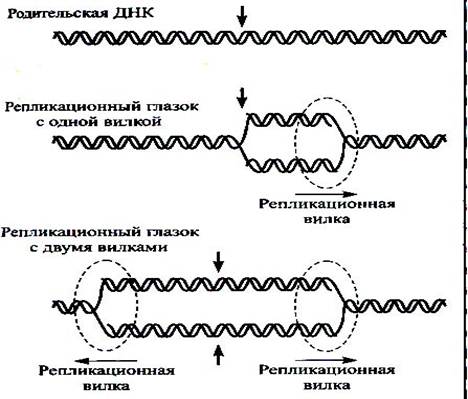
Рис.2.Возможное движение реплткативной вилки.