 2015-04-01
2015-04-01 4118
4118Согласно общепринятым представлениям, в основе поведения животных в сообществе лежит взаимодействие двух факторов: 1) общественного инстинкта, побуждающего их объединяться с сородичами и поддерживать с ними постоянные контакты; 2) внутривидовой агрессивности, которая позволяет установить и поддерживать определенный порядок в сообществе. На значение третьего фактора – уровня развития рассудочной деятельности животных данного вида, который определяет сложность отношений в сообществах высокоорганизованных млекопитающих и птиц, – одним из первых указал Л.В. Крушинский (1973) (см.: 5.3.9). Впоследствии о значении этого фактора в установлении "неформальных контактов" между животными писали неоднократно.
5.3.1. ИЕРАРХИЯ ДОМИНИРОВАНИЯ
Структурированность сообщества – поддержание иерархической организации, обеспечивается прежде всего благодаря феномену доминирования и подчинения.
Норвежский эколог Т. Шьелдерупп-Эббе (Schelderupp-Ebbe, 1924) обнаружил в группах домашних кур и уток строгую упорядоченность отношений между птицами. Каждая особь либо превосходит по силе партнера, либо уступает ему. Эти взаимоотношения были названы "порядком клевания". При формировании группы происходит "выяснение отношений" птиц друг с другом, в ходе которого постепенно выделяется одна, которая первой получает доступ к корму и гоняет от него всех остальных. Ниже ее на "лестнице доминирования" располагается птица второго ранга, которая превосходит всех, кроме главной, доминантной особи, и так далее. В самом основании находится особь, которую гоняют все члены группы.
Такая иерархическая система вырабатывается при столкновении птиц в борьбе за "ограниченный ресурс" (за место на насесте, пищу), и на ранних этапах ее установления происходит много драк. Однако когда иерархия устанавливается, она оказывается стабильной, поскольку порядок соподчинения особей устойчиво поддерживается. Обычно при приближении высокоранговой птицы подчиненные особи уступают ей без сопротивления.
Анализ показал, что предпосылкой стабильности иерархии является индивидуальное узнавание особей. В экспериментах, в которых одну и ту же птицу помещали последовательно в разные группы, "откалиброванные" таким образом, что она занимала в них разное иерархическое положение, куры обнаружили незаурядные способности к запоминанию и распознаванию членов каждой группы и без колебаний занимали причитающееся им положение.
У кур иногда устанавливается совершенная линейная иерархия, так что ни одна птица никогда не клюет особей, стоящих выше ее на иерархической лестнице (табл. 5.1). Подобные "идеальные" сообщества исключительно редки. Среди беспозвоночных их образуют, например, сверчки и речные раки, у которых иерархические отношения также строятся на основе индивидуального распознавания. В то же время у большинства видов животных обнаруживаются различные отклонения ац строгого линейного порядка.
Описанное явление – формирование иерархической структуры группы – стало рассматриваться как механизм, благодаря которому одно или несколько животных получают приоритет во всех жизненных ситуациях.
Предполагалось, что иерархическое ранжирование выделяет наиболее жизнеспособных особей, обеспечивая преимущественный успех их потомства в процессе естественного отбора.
Таблица 5 1
"Идеальная" линия иерархии в группе, состоящей из 12 кур
Примечание. В опыте каждая из птиц была индивидуально помечена. Таблица составлена на основе этограмм регистрации контактов птиц между собой в течение определенного времени наблюдения В вертикальных столбцах – число клеваний, которые данная курица нанесла другим членам группы, в горизонтальных – число клеваний, которые она же получила от других членов группы
"Идеальность" данной иерархической структуры выражается в том, что ни одна курица не клюнула ни одну из особей, стоящих на иерархической лестнице выше ее
| Y | в | V | R | G | YY | ВВ | W | RR | GG | YB | BR | |
| Y | ||||||||||||
| В | 22 | |||||||||||
| V | S | 29 | ||||||||||
| R | 18 | 11 | 6 | |||||||||
| G | 11 | 21 | 11 | 12 | ||||||||
| YY | 30 | 1 | 6 | 21 | 8 | |||||||
| ВВ | 10 | 12 | 3 | 8 | 15 | 30 | ||||||
| W | 12 | 17 | 21 | 6 | 3 | 19 | 8 | |||||
| RK | 17 | 26 | 12\ 11 | 10 | 17 | 3 | 13 | |||||
| GG | 6 | 16 | 7 | 26 | 8 | 6 | 12 | 26 | 6 | |||
| YB | 11 | 7 | 2 | 17 | 12 | 13 | 11 | 18 | 8 | 21 | ||
| BR | 21 | 6 | 16 | 3 | 15 | 8 | 12 | 20 | 12 | 6 | 27 |
Действитель но, некоторые эксперименты прямо свидетельствовали о лучшей приспособленности членов жестко организованных сообществ. Например, в одних группах кур доминантную особь регулярно удаляли и заменяли незнакомой птицей, так что остальные вынуждены были постоянно заниматься "выяснением отношений", тогда как членов контрольных групп не тревожили. В контрольных группах агрессивные стычки происходили реже, яйценоскость кур была выше, т.е. группы с постоянным составом обладали явным преимуществом благодаря стабильной социальной среде. Аналогичную иерархическую структуру и свойства' имеют колонии серых крыс (см.: 5.5.1).
 Один из наиболее распространенных методических приемов, в особенности при исследовании физиологических основ доминирования и агрессивности, – это экспериментальный анализ поведения в "конкурентных ситуациях", когда моделируется ограниченность доступа к ресурсам. Для этого двум животным, на определенное время лишенным пищи или воды, одновременно открывают путь к одной кормушке или поилке. В зависимости от условий эксперимента подкрепление может получить либо только один из конкурентов, либо доминантом считается животное, которое больше съедает или дольше контролирует кормушку.
Один из наиболее распространенных методических приемов, в особенности при исследовании физиологических основ доминирования и агрессивности, – это экспериментальный анализ поведения в "конкурентных ситуациях", когда моделируется ограниченность доступа к ресурсам. Для этого двум животным, на определенное время лишенным пищи или воды, одновременно открывают путь к одной кормушке или поилке. В зависимости от условий эксперимента подкрепление может получить либо только один из конкурентов, либо доминантом считается животное, которое больше съедает или дольше контролирует кормушку.
Эксперименты такого рода, проведенные на животных разных видов, показали, что корреляция между разными показателями доминирования (конкуренция за пищу, воду, территорию, доступ к половому партнеру, возможность уйти в гнездо и т.п.) может быть слабой или совсем отсутствовать. Даже в строго контролируемых лабораторных условиях при использовании генетически однородных однополых и одновозрастных животных не найдено "единого" доминанта.
5. 3.2. РОЛЬ АГРЕССИИ В ПОДДЕРЖАНИИ СТРУКТУРЫ СООБЩЕСТВА
Образование и поддержание стабильных общественных группировок невозможно без внутривидовой агрессии (см.: рис. 5.2). Она выполняет следующие функции:
1)  обеспечение изоляции группы в пределах одной популяции, что, например, препятствует перемешиванию стай при встрече их на одной территории;
обеспечение изоляции группы в пределах одной популяции, что, например, препятствует перемешиванию стай при встрече их на одной территории;
2) противодействие проникновению в данное сообщество чужих особей, а также ограничение числа производителей путем изгна ния части самцов; в таких случаях возникает "резерв" мигрирую щих особей, за счет которого пополняется недостаток производи телей в других популяциях; у многих видов существование подоб ного резерва обеспечивает также обмен генами между группами;
3) участие в формировании и поддержании структуры индивидуали зированных сообществ.
Иерархические отношения между животными устанавливаются в результате конфликтов – либо настоящих драк (рис. 5.3), либо угрожающих демонстраций, причем последние бывают не менее эффективными, чем первые (рис. 5.4). Как правило, они включают различные приемы, благодаря которым животные кажутся крупнее, чем они есть на самом деле. Например, хождение на двух ногах, вздыбливание шерсти, подскакивание шимпанзе действуют не менее эффективно, чем прямое нападение. Такие же элементы поведения обнаруживаются у ящериц (рис. 5.4). Птицы, демонстрируя силу, распушают перья и расправляют крылья.
К. Лоренц предположил, что существование групп с тесными индивидуальными связями между особями возможно только у животных с достаточно развитой способностью к направленной агрессии, у которых объединение двух или большего числа особей имело значение для выживания. Этой проблеме посвящена его книга "Агрессия (так называемое зло)", написанная в 60-е годы и переведенная на русский язык в 1994 г.
Логический анализ наблюдений за поведением животных при проявлениях агрессивности, страха или взаимного влечения позволил Лоренцу сделать заключение о существовании тонкого баланса между этими состояниями, а также о неотделимости проявлений агрессивности как центробежной тенденции, от проявлений "любви" – общественного инстинкта, которые обуславливают взаимное притяжение особей.
Написанная очень увлекательно, книга Лоренца не только внесла вклад в специальную область биологии – этологию, но и явилась одним из важнейших научно-философских произведений XX в. В ней приведены многочисленные доказательства того, что агрессивность является врожденным, наследственно обусловленным свойством всех высших животных. Прослеживая подобные свойства в поведении человека, К. Лоренц приходит к выводу, что "есть веские основания считать внутривидовую агрессию наиболее серьезной опасностью, какая грозит человечеству в современных условиях культурно-исторического и технического развития" (Лоренц, 1994). Это положение повлекло за собой обвинения автора в недооценке социальной природы человека. Оно способствовало отрицательному отношению к этой книге в СССР и на десятилетия задержало ее перевод на русский язык. Между тем, основное значение книги заключается прежде всего в изложении основ регуляции социальных отношений у животных.
5.3.3. РИТУАЛЫ И ДЕМОНСТРАЦИИ
Поддержание структуры сообщества осуществляется на основе агонистического поведения, т.е. комплекса реакций нападения, угрозы, подчинения и бегства. Как уже упоминалось, установление иерархии может происходить в результате прямых столкновений и драк, тогда как по мере ее упрочения в большинстве случаев прямые проявления агрессии уступают место угрозам и демонстрациям.
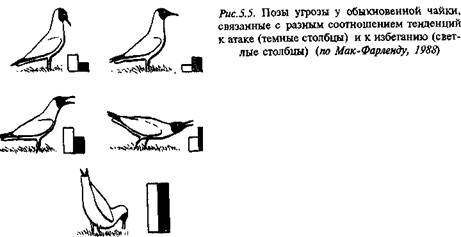
Выше (раздел 4.3) были рассмотрены типичные примеры угрожающих демонстраций у рыб и чаек. Согласно гипотезе Н. Тинберге-на, выраженность практически всех демонстраций определяется соотношением уровня агрессивности и страха, которые в данный момент переживает животное (рис. 5.5 и 5.11).
 Знаменитая серия зарисовок немецкого этолога П. Лейхаузена показывает мимику домашней кошки при разных сочетаниях уровней агрессии и страха (рис. 5.6): агрессивная тенденция усиливается слева направо, страха – сверху вниз.
Знаменитая серия зарисовок немецкого этолога П. Лейхаузена показывает мимику домашней кошки при разных сочетаниях уровней агрессии и страха (рис. 5.6): агрессивная тенденция усиливается слева направо, страха – сверху вниз.
Другую группу демонстраций составляют ритуализованные движения, тормозящие агрессию со стороны более сильных сородичей. Их называют позами "покорности" или "умиротворения". По всем своим параметрам они противоположны жестам угрозы. Так, коралловая рыбка, угрожающая сородичу, демонстрирует ему свой яркий наряд, распахивает плавники или жаберные крышки, движется резко, демонстрируя силу. Когда же она "просит пощады", все выглядит совершенно по-другому: ее окраска бледнеет, она прижимает плавники, поворачивается к сопернику узким сечением тела, стараясь казаться маленькой, движется медленно, крадучись. Подобным же образом, демонстрируя угрозу, птицы распушают перья, чтобы казаться как можно крупнее, тогда как в знак подчинения они прижимают перо, и их силует становится более тонким. У макаков-резусов позы и движения доминантного и подчиненного животных диаметрально различны (рис. 5.7 и 5.11).

Выразительные позы угрозы и подчинения у воронов (рис. 5.8) не оставляют сомнений в их функциональном "назначении".
Формы ритуалов умиротворения многообразны, тем не менее некоторые из них (поза подставления, реакция выпрашивания корма) у самых разных групп животных одинаковы (рис. 5.9).
К. Лоренц (1994) выделил несколько основных особенностей таких ритуалов.
1. Демонстративное подставление наиболее уязвимой части тела. Так, побежденный волк отворачивает голову и подставляет победителю чрезвычайно ранимую боковую сторону шеи, выгнутую навстречу укусу. Галка подставляет под клюв птицы, которую нужно умиротворить, свой незащищенный затылок – обычную мишень при серьезном нападении с целью убийства. У многих видов птиц оперение на таких участках тела имеет особую окраску. У галок оно шелковисто-серое, а у техврановых, которые крупнее и темнее на затылке имеется более светлое пятно. По данным Н. Тинбергена (1974), чайки, демонстрируя умиротворение, отворачивают от противника клюв (см.: рис. 4.6 в), при этом привлекает внимание их белоснежный затылок (как у обыкновенной чайки) или особый рисунок на светлом фоне (как у молодой моевки).

 2. Воспроизведение некоторых элементов детского поведения. Распространенный вариант ритуала умиротворения у птиц - это имитация позы птенца, выпрашивающего корм (рис. 5.10).
2. Воспроизведение некоторых элементов детского поведения. Распространенный вариант ритуала умиротворения у птиц - это имитация позы птенца, выпрашивающего корм (рис. 5.10).
В репертуаре социального поведения псовых жесты умиротворения также играют важную роль; они происходят непосредственно из отношений щенка с его матерью и являются производными от движений сосания или просьбы накормить (см.: рис. 5.9). Как предполагает К. Лоренц (1994), эти действия превратились в ритуалы у тех животных, у которых изначально существовали более общий, социальный смысл, оказалось возможным формирование крупных социальных групп на базе супружеских пар и семей.


 |
3. Выражение социальной покорности с помощью действий, характерных для поведения самки при спаривании. Эти действия типичны для многих обезьян, особенно павианов. Ритуальный поворот задней части тела означает признание более высокого ранга другой обезьяны. Позу подставления они выполняют с самого раннего возраста без всякого обучения, даже при воспитании в изоляции от сородичей (рис. 5.12). Как уже упоминалось, те части тела, которые животные демонстрируют в знак угрозы или подчинения (рели-зеры – см.: 3.4), окрашены особенно ярко и заметно, чем подчеркивается выразительность церемонии. Так, у некоторых видов павианов задняя часть тела, которую они демонстрируют противнику в позе подставления, раскрашена самым фантастическим образом.
5.3.4. СЛОЖНЫЕ СИСТЕМЫ ИЕРАРХИИ
Представление о том, что в условиях ограниченного доступа к какому-либо ресурсу доминирование способствует его получению в первую очередь, подтверждено в многочисленных исследованиях социального поведения животных (подробнее см.: Резникова, 1997).
Сростом числа исследований на животных разных видов выявилось множество отклонений от классической линейной схемы иерархической структуры сообществ. Наиболее распространенное отклонение – превращение "лестницы" или "цепочки" в двухуровневую структуру: доминант – и все остальные. Случаи такого абсолютного доминирования наблюдаются, например, при содержании самцов домовых мышей или самцов колюшки в ограниченном пространстве. В иных случаях в иерархических "цепочках" могут наблюдаться "петли" или "треугольники", когда особь А гоняет особь В, особь В – особь С, но А боится С. Описаны даже такие ситуации, когда длинная иерархическая цепочка замыкалась в круг.
Показано, что в сообществах могут действовать параллельные системы иерархии – у самцов и у самок. Изменение ранга самки, как это было описано К. Лоренцем (1978) при изучении сообщества галок, может происходить, если она становится супругой высокорангового самца. У большинства видов самцы имеют более высокие ранги, однако имеются и исключения.
У некоторых видов животных внутри большого сообщества существуют стабильные группировки – кланы, чаще всего образованные одной или несколькими самками с их потомством одного-двух поколений (см.: Крук, 1977). Такие "материнские кланы" можно наблюдать, например, у слонов (И. и О. Дуглас-Гамильтон, 1981), дельфинов (Прайер, 1981; см. также обзор: Крушинская, Лисицына, 1983), гиеновыхсобак (Цж. и Г. Лавик-Гудолл, 1977). На рис. 5.13 приведена схема "построения" труппы павианов при перемещении на открытом пространстве.

Помимо относительно стабильных группировок и кланов, члены сообщества могут образовывать и временные коалиции, направленные либо на достижение "сиюминутной выгоды", либо на то, чтобы повысить свой общий ранг. Множество примеров подобных ситуаций дают исследования приматов (японских макаков, зеленых мартышек, павианов, шимпанзе). Так, два–три самца могут объединиться, чтобы отвлечь доминанта, инсценируя драку или интересную находку, и, пока доминант "наводит порядок", по очереди получить доступ к пище или даже увести самку. Пример такой же "эксплуатации" молодым самцом принятого в сообществе доброжелательного отношения ко всем детенышам, можно видеть на рис. 5.14.
Существует достаточно много примеров, когда в сообществе приматов складывается не столько иерархия особей, сколько иерархия группировок (Гудолл, 1992).
5.3.5. ЛАБИЛЬНОСТЬ ИЕРАРХИЧЕСКОЙ СТРУКТУРЫ В ИНДИВИДУАЛИЗИРОВАННЫХ СООБЩЕСТВАХ
Особенности структуры сообществ, выявленные в наблюдениях за животными разных видов, показали, что внутривидовая иерархия, ее структура и механизмы поддержания, а также возможные отклонения зависят от ряда факторов.
Конкретная ситуация в сообществе почти всегда лабильна и также может выходить за рамки простых иерархических отношений. Этологи выделяют некоторые правила, соблюдающиеся в группах животных в обход существующей иерархической системы. Одно из таких правил – так называемое "право обладания". Так, например, если низкоранговая особь шимпанзе добывает редкое лакомство, все остальные окружают ее, выпрашивая свою долю. Высокоранговые животные при этом не пользуются никакими преимуществами.
Пластичность поведения животных в сообществе обусловлена их богатым индивидуальным и социальным опытом. Создается впечатление, что классическая линейная иерархия является лишь некой врожденной "матрицей", характерной для многих общественных животных. В течение жизни на нее накладывается приобретенное умение оценивать и использовать социальные связи.
Это положение было подтверждено экспериментально (Anderson, Mason, 1978). Были сформированы две группы макаков-резусов: одна состояла из обезьян, выращенных в изоляции, с "искусственными" матерями (как в известных опытах Harlow et al., 1963), а члены другой росли со своими матерями и сверстниками. В обеих группах экспериментаторы искусственно создавали конкуренцию за бутылочку с водой. Оказалось, что у животных, выращенных в изоляции, устанавливается стабильный порядок доступа к бутылочке, напоминающий "порядок клевания" у кур. В то же время у животных с нормальным опытом социальных контактов была обнаружена достаточно высокая лабильность социального поведения. Они были, например, способны объединяться в коалиции, могли искать защиты у доминанта в спровоцированной ими же драке и тем самым отвлечь его от бутылочки и т.п. Подобные закономерности обнаружены и у беспозвоночных, например в опытах В.А. Харькива (1993) на муравьях (подробнее см.: Резникова, 1997).
Форма общественной организации может варьировать у данного вида в зависимости от местообитания и плотности популяции. Существует ряд четких примеров изменения структуры сообщества в зависимости от плотности населения. Часто перенаселение ведет к появлению в сообществе более выраженной иерархии. В экспериментах, проведенных на таких разных животных, как луна-рыба и мышь, было обнаружено, что в условиях высокой плотности самцы образуют типичную иерархию, но когда, позволяет пространство и существуют укрытия, они переходят к территориальности.
Зависимость формы организации сообществ, в частности размера групп, от экологических условий лучше всего исследована у млекопитающих. Однако суммировать весь ряд факторов, влияющих на величину группы и характер внутригрупповых взаимоотношений, далеко не просто. На первый план выступают два фактора: распределение пищевых ресурсов и давление со стороны хищников.
Если ресурсов питания много, то это способствует образованию больших групп. Например, у пасущихся на равнинах травоядных большие группы мало угрожают сохранению пищевых ресурсов. Напротив, виды, находящиеся на вершине пищевой цепи (такие как волк или крупные кошки), не могут существовать при высокой плотности.
Что касается второго фактора – угрозы со стороны хищников, то очевидно, что большим группам справляться с этой угрозой легче.
Демографические параметры популяции, влияющие на характер взаимоотношений особей в группах, зависят от обоих этих фак/ор- ов. Это относится в первую очередь к числу взрослых самцов в группах: в областях с большим давлением со стороны хищников оно заметно выше (подробнее см.: Резникова, 1997). I
Одно из проявлений пластичности структуры сообществ животных – ее изменения в зависимости от степени урбанизации среды. Воздействие антропогенных факторов может существенно влиять на пищевые ресурсы и наличие мест для гнездования птиц, меняя соответствующим образом размер и способ организации сообществ. Ярким примером таких влияний могут быть изменения, которые претерпели сообщества многих видов врановых птиц. Например, галки (Corvus monedula), типично колониальный вид, могут селиться разреженно и даже образовывать колонии ленточного типа. Напротив, представители одиночно гнездящегося вида – серые вороны (Corvus comix) – в городах при изобилии корма и гнездового материала строят гнезда в непосредственной близости друг от друга, а в единичных случаях – даже на одном дереве (Константинов, 1992).
Говоря о пластичности социального поведения птиц, нельзя особо не упомянуть о вброне (Corvus corax., L.). Ворон считался чисто одиночным видом, однако сейчас показано, что вороны могут сотнями кормиться на свалках и других массовых источниках корма (Константинов, 1992). Природа таких стай может быть очень сложной, и их нельзя считать простыми скоплениями в указанном выше смысле (см.: 5.2.2). Американский зоолог Б. Хейнрих(1994) в течение многих лет наблюдал за поведением воронов в лесах штата Мэн. Приманками для них служили коровьи туши или специально разложенные куски мяса. Стаи воронов (он назвал их "ордами"), насчитывавшие от 15 до 300 птиц, постепенно расклевывали эти приманки. Наблюдения позволили установить, что такая концентрация воронов у приманок – явление совершенно особое; эти объединения отличны по своей природе от стай других врановых, которые кормились тут же. В частности, голубые сойки (Cyanocitta cris-tata, L.) применяют альтернативную стратегию и, обнаружив корм, стараются скрыть это, чтобы не делиться с сородичами.
Самая главная особенность "орд" воронов состоит в том, что это не пассивные скопления. В поисках корма они летают парами или поодиночке и, обнаружив его, специальными криками ("воплями") широко извещают об этом, как бы приглашая всех желающих присоединиться. Эти крики нельзя услышать практически ни в какой другой ситуации. Хейнрих предполагает, что они развились из звуков, которые издают птенцы, выпрашивая корм.
Детальный анализ, который мы не будем приводить подробно, показал, что "приглашающие вопли" издают только молодые птицы. Благодаря этому они получают или удерживают доступ к корму, до которого их иначе не допустили бы владельцы территории – взрослая пара воронов. Хейнрих предполагает, что, помимо обеспечения пищей, у этих сборищ молодых птиц есть и другое предназначение – обретение социального статуса и "помолвки". Bopok, пока не имеющий пары, найдя корм, приглашает других птиц присоединиться к нему и тем самым не только получает или сохраняет доступ к пище, но, возможно, повышает свой статус, демонстрируя способности будущего кормильца при выведении потомства.
5.3.6. ДОМИНИРОВАНИЕ И РЕПРОДУКТИВНЫЙ УСПЕХ
Этологические и генетические исследования показали, что в сообществах мышей иерархическая структура лабораторной популяции определяет преимущественное отцовство доминанта (McCleam, DeFries, 1972).
Однако, по данным исследований индивидуализированных сообществ высокоразвитых животных, связь между иерархическим рангом и числом оставленных потомков бывает далеко не очевидной. Подчиненная особь может успешно адаптироваться к поведению доминанта и получать "все блага жизни", даже не вступая в борьбу. У самца благородного оленя описана удивительная "клепто-реп-родуктивная стратегия". Молодой самец успешно спаривался с самками, пока хозяин "гарема" отвечал на вызовы других самцов.
5.3.7. ИЕРАРХИЯ РОЛЕЙ И «РАЗДЕЛЕНИЕ ТРУДА" В СОЦИАЛЬНЫХ ГРУППИРОВКАХ ЖИВОТНЫХ
Усложнение схемы иерархического строения сообщества связано и с "распределением ролей" (или "разделением труда") животных в группе. Оно описано у некоторых совместно охотящихся хищных рыб (тунца, макрели), а также у ряда видов млекопитающих, таких как бобры, львы, волки, гиены, гиеновые собаки, шакалы и др. Этот феномен представляет собой выполнение членами группы различных, но четко определенных по функции действий, например при охоте или охране территории. Возможность выполнения разных ролей в сообществе определяется у каждой особи сложнейшим сочетанием видовых, наследственных ФКД и поведения, основанного на индивидуальном и социальном опыте. На "результирующей" влияния опыта и врожденных задатков и основана роль каждого индивидуума в сообществе.
В целом, "разделение труда" в сообществах животных, несомненно, делает их социальные отношения более сложными и многообразными. В разных ситуациях на первый план могут выходить особи, более способные к тому или иному виду деятельности. Такие отношения называют "ролевой иерархией".
Замечательное разделение труда существует в сообществе бобров. Обитающая в хатке группа животных выделяет "дежурных", которые по очереди следят за бобрятами. Они постоянно играют роль "спасателей", так как детеныши еще не очень хорошо плавают и, покидая хатку, могут не найти входа в нее, задохнуться в воде либо погибнуть на берегу. Бобры, работающие вне хатки, также вь^пол-няют разные функции, такие как снабжение бобрят пищей, охрана или строительство.
Существуют наблюдения, свидетельствующие о том, что у некоторых видов "разделение труда" включает в себя манипуляции поведением одних особей со стороны других. Наиболее известны, хотя и не полностью убедительны, знаменитые эксперименты К. Моуэра (Mower, 1940): в камеру помещались несколько крыс, которые могли нажимать на рычаг для получения пищи, однако кормушка находилась в отдалении от него. Оказалось, что в такой ситуации на рычаг нажимали лишь немногие особи, обеспечивая пищей всех остальных. Остается неясным, почему "работала" только часть животных, а другие вели "паразитический" образ жизни.
 Другой, более убедительный пример "разделения труда" и "эксплуатации" в группе молодых шимпанзе описал Л.А. Фирсов (1977). Он провел свои наблюдения на озерном острове в Псковской обл., где молодые животные летом вели свободный образ жизни. На первом этапе эксперимента шимпанзе могли получить приманку из открывающегося ящика только в результате успешных действий партнера. Когда они усвоили этот навык, отношения между ними начали изменяться. Достаточно сбалансированные в начале эксперимента, они быстро переходили в фазу "беззастенчивой эксплуатации", когда большую часть заработанной приманки получали не "работники", а "наблюдатели". Прежняя картина отношений восстанавливалась лишь после энергичных столкновений. Несомненно, такое "разделение труда", особенно путем манипуляций поведением партнеров, требует известного социального опыта и гибкости поведения.
Другой, более убедительный пример "разделения труда" и "эксплуатации" в группе молодых шимпанзе описал Л.А. Фирсов (1977). Он провел свои наблюдения на озерном острове в Псковской обл., где молодые животные летом вели свободный образ жизни. На первом этапе эксперимента шимпанзе могли получить приманку из открывающегося ящика только в результате успешных действий партнера. Когда они усвоили этот навык, отношения между ними начали изменяться. Достаточно сбалансированные в начале эксперимента, они быстро переходили в фазу "беззастенчивой эксплуатации", когда большую часть заработанной приманки получали не "работники", а "наблюдатели". Прежняя картина отношений восстанавливалась лишь после энергичных столкновений. Несомненно, такое "разделение труда", особенно путем манипуляций поведением партнеров, требует известного социального опыта и гибкости поведения.
Сходные отношения складывались и в группе молодых ворон (в начале эксперимента их возраст составлял 2 месяца), которые обучались нажимать на рычаг для получения корма одновременно. И в этом случае доля пищедобывательных реакций, совершенных членами группы, с самого начала была различной, и это различие углублялось в процессе эксперимента. В конце концов около 80% всех реакций совершала одна из птиц, получая при этом лишь 38% заработанных ею порций корма. Остальные 3 птицы получали в среднем по 20% всех подкреплений, даже если совершали не более 2% нажатий на рычаг. Существенно отметить, что взаимодействия между птицами были почти лишены признаков агрессивности, во всяком случае, ворона, нажимавшая на рычаг, не пыталась отгонять других членов группы от "заработанного" ею корма. Возможно, это происходило потому, что самыми активными "добытчиками" были "низкоранговые" особи (Зорина, 1996). Таким образом, в данной ситуации представители весьма разных групп позвоночных ведут себя довольно сходным образом. Изучение индивидуализированных сообществ животных дает много примеров крайне важной
роли сотрудничества для
успешной деятельности и выживания группы. Наиболее яркие из них – это совместная охота у многих крупных хищников, совместное сложное строительство и выращивание потомства у далеких друг от друга видов, например у бобров, голых землекопов (см.: 5.5.1) и общественных насекомых – термитов, ос, пчел, муравьев (см.: 5.6.5).
В приведенных примерах сотрудничество между членами сообщества составляет видоспецифическую черту их поведения, непременно характерную для всех особей. Наряду с этим сотрудничество может возникать как индивидуальное приспособление отдельных особей к конкретным условиям среды, которое не предусмотрено видовыми стереотипами поведения. Такой вид сотрудничества представляет особый интерес для характеристики разумных компонентов в поведении животных (см.: Крушинский, 1986).
Наиболее убедительные примеры этих высших форм кооперации обнаружены у человекообразных обезьян. Так, Дж. Гудолл (1992) приводит в своей книге ряд примеров, когда шимпанзе оказывали помощь другим членам группы, которые.могли и не быть их родственниками. Еще один пример – наблюдение Р. Футса за "говорящей" обезьяной Уошо (см. также: 5.4.2.2), которая жила на островке, окруженном рвом с водой и изгородью с электрическим током. Однажды молодая самка Синди, решив навестить Уошо, каким-то образом преодолела эту изгородь, но упала в воду и начала тонуть. Увидев это, Уошо, держась за траву, вошла в воду и умудрилась схватить Синди за руку, когда та, может быть, в последний раз всплыла на поверхность. Следует подчеркнуть, что между обезьянами не было родственных отношений, да и познакомились они совсем незадолго до события (рис. 5.15).
Разбирая этот и подобные довольно многочисленные примеры, Дж Гудолл полагает, что отбор родичей может играть роль в формировании способности к взаимопомощи не только среди родственных, но также и посторонних друг другу особей.
О способности к сотрудничеству не только у приматов, но и других высокоорганизованных позвоночных свидетельствуют упомянутые выше данные о врановых птицах. Своеобразным экспериментальным доказательством ее существования могут служить также данные опытов Л. С. Бондарчука, описанные в работе Крушинского и др. (1982), по одновременному обучению двух ворон в специальной камере, разделенной пополам прозрачной перегородкой. В каждой половине находилась педаль, при нажатии на которую появлялся корм, но это происходило только тогда, когда вороны нажимали на "свои" педали одновременно. Независимо от этого ворон научили открывать дверцы примыкавших к обоим отделениям камеры дополнительных клеток, где птицы видели кормушку с мясом. Каждый из навыков вырабатывался достаточно легко. Затем в главном эксперименте одну из ворон запирали в клетку, так что вторая птица при нажатии на педаль корма не получала.
Оставшись в одиночестве, ворона в конце концов открывала клетку и выпускала партнера. Этот факт рассматривается как доказательство способности птиц к экстренной интеграции независимых навыков в новой ситуации, т.е. к одному из относительно простых видов рассудочной деятельности (Зорина, 1997).
Анализируя проблему взаимопомощи и сотрудничества у высших позвоночных, Л. В. Крушинский (1986) приходит к выводу, что основным механизмом, обеспечивающим возможность таких взаимоотношений между членами сообщества, может быть только достаточно развитая рассудочная деятельность. Действительно, поведение Уошо в приведенном примере включает экстренную оценку новой, ранее никогда не имевшей места ситуации, способность поставить себя на место обезьяны, терпящей бедствие, и принять адекватное данной ситуации решение.
5.3.8. РАЗВИТИЕ СОЦИАЛЬНОГО ПОВЕДЕНИЯ В ОНТОГЕНЕЗЕ
Развитие социального поведения животных в онтогенезе зависит, по крайней мере, от двух основных факторов. Первый из них – это генетически детерминированная видоспецифическая "система общения", т.е. система коммуникации с особями своего вида, основанная на обмене сигнальными позами, жестами, движениями, включая демонстрацию особых сигнальных структур релизеров (см.: 3.4). Как само продуцирование подобных сигналов, составляющих язык животных, так и их опознание, как правило, не требуют индивидуального опыта: оно относится к категории врожденных реакций. Второй фактор – существование определенных периодов индивидуального развития, в течение которых с наибольшей легкостью и быстротой формируются связи между особями: запечатле-ние детьми внешнего облика родителей, которое будет определять общественные взаимодействия этих особей по достижении ими взрослого состояния (см. также: 3.5 и 7.1).
Важнейшая роль в становлении социального поведения (как и практически любых других его форм) принадлежит играм (см. обзор: Зорина, 1998). Особенно ярко роль игры проявляется у животных, которые живут в сложно организованных сообществах. В ходе игры используются элементы агонистического поведения и закладываются основы иерархических отношений между их участниками. По мере взросления игры многих животных приобретают все более грубый характер и нередко заканчиваются агрессивными эпизодами. Благодаря этому животные не только получают сведения о сильных и слабых сторонах своих партнеров по игре и об относительном иерархическом положении своей матери и матерей товарищей по играм, но и учатся драться, угрожать, устанавливать союзнические отношения. Это позволяет им впоследствии успешно конкурировать с другими членами сообщества, в котором способность отстоять свои права и повысить ранг часто зависит от умения драться.
Установлено, что в характере социальных игр четко отражается видовая специфика поведения взрослых животных. Как отмечает К.Э. Фабри (1993), у одних видов грызунов (морские свинки) игровая борьба отсутствует, а социальные игры сводятся к сигналам "приглашения". В отличие от них, у большинства других грызунов распространены именно игровые схватки. Особенно отчетливо игровые взаимодействия грызунов проявляются в физически сложной среде обитания (Мешкова, Федорович, 1996).
Социальные игры очень характерны для хищных млекопитающих. В качестве примера современных исследований этого аспекта проблемы можно привести данные многолетних наблюдений Н.Г. Овсянникова (1993) за поведением и социальной организацией песцов (Alopex galopus, L). Они свидетельствуют, что взаимодействия молодых песцов в процессе игры действительно обеспечивают механизмы социальной интеграции в выводках этих животных. Игровая борьба у песцов феноменологически не имеет ничего общего с настоящей агрессией, хотя отдельные движения могут быть похожими. В целом схватки зверьков в процессе игры производят впечатление более стереотипных, монотонных действий, чем при настоящих схватках. Автор приводит ряд доказательств того, что игровая борьба эмоционально положительна и оказывает на выводки интегрирующее влияние. По свидетельству Овсянникова, во время игры стираются различия социального положения и роли в сообществе, временно ослабевает психосоциальный стресс, который неизбежен при взаимодействиях по необходимости – для выращивания потомства, добывания пищи и т.п.
Соотношение игровой борьбы, подвижных и охотничьих игр у разных видов также различно. При этом, как отмечает К.Э. Фабри (1993), нужно учитывать, что сами по себе элементы социального поведения представляют собой ФКД, которые проявляются в "готовом" виде и не требуют тренировки для своего созревания. Роль игры в становлении социального поведения состоит в том, что, если на ранних этапах она складывается из отдельных компонентов, то по мере взросления эти компоненты все больше интегрируются в единое целое.
Один из вариантов социальных игр – игры матери с детенышем. Они характерны для хищных млекопитающих, но особенно развиты и выражены у человекообразных обезьян: мать играет с детенышем с первых же месяцев жизни и до окончания подросткового периода.
В процессе онтогенеза у многих животных меняются образ жизни и соотношение разных типов контактов. Так, например, молодые лисицы и медведи живут семейными группами, в которых проявляются многообразие и сложность дружелюбных взаимодействий, а став взрослыми, ведут одиночный образ жизни. При этом у взрослых бурых медведей дружелюбные контакты, свойственные молодым, полностью уступают место выраженной агрессивности по отношению друг другу (Пажетнов, 1985; 1991).
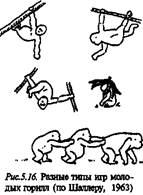
Наиболее сложный процесс развития проходит социальное поведение человекообразных обезьян. На рис. 5.16 показано, как могут играть молодые гориллы.
По наблюдениям Гудолл (1992), детеныши шимпанзе имеют достаточно долгий подростковый период, когда в тесных контактах с сородичами они овладевают не только многочисленными пищедо-бывательными и оборонительными навыками, но усваивают и основные паттерны коммуникативных реакций, и правила поведения в сообществе.
5.3.9. ВЛИЯНИЕ УРОВНЯ РАЗВИТИЯ ЭЛЕМЕНТАРНОЙ РАССУДОЧНОЙ ДЕЯТЕЛЬНОСТИ НА СПЕЦИФИКУ ОБЩЕСТВЕННЫХ ОТНОШЕНИЙ ЖИВОТНЫХ
Л.В. Крушинский (1973) одним из первых систематизировал данные о том, что степень многообразия и пластичности отношений в сообществе животных тесно связана с уровнем их психического развития. В своей монографии "Биологические основы рассудочной деятельности" он рассмотрел специфику общественных отношений у животных, обладающих высокоразвитой рассудочной деятельностью, – врановых, дельфинов, хищных млекопитающих (псовых, кошачьих, медведей) и разных видов приматов, сопоставив ее с уровнем развития их рассудочной деятельности по экспериментальным данным и по наблюдениям в природе (Крушинский, 1986. С. 176-208).
Ниже (см.: 5.4) рассмотрена организация сообществ этих животных в других аспектах, не дублирующих содержание упомянутого раздела книги Крушинского, к которой мы и отсылаем читателя.
Л.В. Крушинский пришел к выводу, что в сообществах высокоорганизованных животных взаимопомощь и сотрудничество играют весьма существенную роль, даже более значительную, чем агрессивность, связанная с поддержанием иерархической структуры. Речь идет не о том, что этот фактор определяет тип организации сообщества: независимо от уровня развития рассудочной деятельности виды могут вести как групповой, так и одиночный образ жизни. На основе приведенных данных Л.В. Крушинский (1986) делает вывод о том, что у наиболее умных животных из рассмотренных им групп – обезьян и дельфинов – наблюдаются и наиболее сложные формы социальных взаимоотношений (см. также: 5.5.3). Они не укладываются в рамки простых территориально-иерархических отношений, формирующихся на основе агрессии. У хищных млекопитающих, по его оценкам характеризующихся более низким уровнем рассудочной деятельности, социальные связи также относительно менее сложны (см.: 5.5.2).
Наиболее полные и убедительные конкретные доводы в пользу такого представления приводит Дж. Гудолл (1992. С. 427–428). Обобщая огромный объем собственных наблюдений за социальными взаимодействиями шимпанзе, она пишет, что это именно та сфера приспособительной деятельности животного, в которой требуются хорошее понимание причинно-следственных связей, мобилизация всех самых сложных познавательных способностей для достижения успеха и поддержания своего социального положения. Так, при рассмотрении вопроса о возрастных изменениях иерархического статуса самцов она указывает, что в ряде случаев борьба за доминирование напоминает "состязание характеров, в котором большое значение имеют... изобретательность и упорство". В ее работе приводятся многочисленные примеры того, как низкоранговая особь достигает желаемой цели с помощью хитроумных обходных маневров, даже при явном неодобрении старшего по рангу. Это требует умения планировать свои действия и манипулировать поведением сородичей (см. также: 5.5.3), что как раз и относится к сфере разумного поведения.
Классическим примером роли такой сообразительности в достижении доминантного положения стал самец Майк, который догадался использовать пустые металлические канистры для усиления эффекта своего угрожающего поведения (Гудолл, 1992). Шимпанзе учитывают тот факт, что положение в группе может улучшиться, если "заключить дружеский союз" с другим самцом.
Соотношение сил в группе шимпанзе постоянно меняется, и каждый из них должен всегда быть настороже, уметь оперативно узнавать особенности сиюминутной ситуации и мгновенно менять в соответствии с ними свое поведение, иначе может последовать суровое возмездие. Как пишет Гудолл, молодой самец, уже направившийся к самке и начавший демонстрировать свои намерения, немедленно остановится и примет нейтральную позу, если на сцене появится самец более высокого ранга.
Наблюдения показывают, что шимпанзе оценивают структуру сообщества отнюдь не только по результатам прямых агрессивных взаимодействий. Благодаря наблюдениям за контактами отдельных особей шимпанзе "вычисляет" на этой основе полную картину и собственное положение в ней: "если А гоняет В, а В угрожает С, следовательно, С ниже рангом, чем А". Практика показывает, что шимпанзе способны к такого рода умозаключениям. Эту сторону поведения некоторые авторы называют "social cognition". Именно подобные факты послужили основанием для высказанного американским исследователем Примэком (Premack, 1983) предположения о том, что у шимпанзе есть такая форма дедуктивного мышления как способность к транзитивному заключению, и побудили изучать ее в физиологическом эксперименте.
Не менее важную роль может играть и присущее шимпанзе умение понимать и оценивать выразительные движения, а также знания, намерения и другие внутренние (умственные) состояния сородичей, строя в соответствии с этим свое поведение. Тесно связанное со способностью к самоосознанию умение мысленно поставить себя на место другого и предугадать его действия Примэк (Premack, 1978; 1983) рассматривал как одно из наиболее сложных проявлений познавательной деятельности животных.
В ряде работ показано, что такой способностью действительно обладают только человекообразные обезьян и. Так, Сифард и Чини (1993), исследуя язык восточно-африканских верветок (см. также: 5.4.3.3), задавались вопросом, могут ли обезьяны понимать разницу между своими собственными представлениями и знаниями и представлениями других особей, а также могут ли они специально проинформировать о чем-то сородича, или изменить неверные представления у другого.
Чтобы ответить на эти вопросы, авторы провели эксперименты с группами макак резусов и японских макак, обеспечивая некоторых особей информацией, которой другие не владели. Особенно красноречивыми оказались результаты экспериментов, в которых мать имела возможность "сообщить" своему детенышу о местоположении корма и о появлении хищника. По условиям опыта мать прекрасно видела, что детеныш не осведомлен, например, о грозящей ему опасности. Оказалось, однако, что у низших обезьян мать никак не пытается воздействовать на поведение детеныша, и, по-видимому, эти животные не принимают в расчет намерения сородичей.
Это вполне соответствует поведению низших обезьян в природе. Например, детеныши верветок, начиная издавать крики тревоги или реагируя на сигналы других, делают много ошибок. Некоторые из них безобидны: например, детеныш издает крик "орел", видя пролетающего над головой голубя. Другие ошибки могут стоить детенышу жизни, например, когда он смотрит вверх, услышав сигнал о приближении змеи. Однако исследователи не обнаружили доказательств того, что взрослые "поправляют" детенышей или как-либо поощряют поведение тех, кто издает сигналы правильно и адекватно на них реагирует. Детеныши верветок учатся только посредством наблюдения, по-видимому, в связи с неспособностью взрослых особей оценить, что уровень знания детенышей уступает их собственному.
Эти результаты перекликаются с данными, полученными Д. По-винелли с соавт. (Povinelly et al., 1992) в остроумном эксперименте с "переменой ролей". Обезьянам (макакам и шимпанзе) предлагали игровой аппарат, напоминающий детский настольный хоккей, в котором партнеры играют попарно и по очереди дергают за рычаги. В данном эксперименте один из двух партнеров (по одну сторону аппарата – животное, по другую – партнер, человек), "информатор", видел, где спрятана приманка, но достать ее не мог (его рычаг был заблокирован), а мог лишь указать на нее второму партнеру – "оператору", который, наоборот, не видел, где находится приманка, и мог ее достать, лишь пользуясь указаниями "информатора" и дергая соответствующий рычаг. Оказалось возможным научить макак хорошо выполнять свои роли: одни животные становились "информаторами", другие – операторами. К перемене ролей они оказались совершенно не способными. Зато шимпанзе прекрасно поняли, что на месте партнера нужно выполнять не свою прежнюю роль, а ту, что на этом месте выполнял человек. Это означает, что шимпанзе, выполняя роль информатора, "влезали в шкуру" оператора, примеривались к его роли и сознавали, что партнер может, а чего не может делать, и наоборот. Поэтому шимпанзе, в отличие от низших обезьян, легко справлялись с задачей перемены ролей. Сходные данные о различиях в степени осознания своих действий и предвидения их результата получены при сравнении орудийного поведения шимпанзе и павианов.
Отрицательные результаты, полученные в опытах с низшими обезьянами, не позволяют выяснить их причину: животное или не способно оценить умственное состояние другой особи, или, сделав такую оценку, оно не может ее использовать.
Продолжая сравнение низших обезьян с шимпанзе, следует упомянуть эксперименты Примэка с шимпанзе Сарой, в которых обезьяна должна была отличить состояние знания и незнания у человека. Ей показывали видеозаписи действий дрессировщиков, пытающихся решить различные бытовые проблемы, например включить кнопкой магнитофон, у которого шнур выдернут из розетки. Во всех случаях Сара указывала возможное верное решение для любимого дрессировщика и... неверное – для нелюбимого. Эти данные доказывают способность шимпанзе поставить себя на место другого и мысленно воспроизвести его действия, а также свидетельствуют о его способности к осознанному обману. Ряд наблюдателей приводят примеры того, как шимпанзе обманывают друг друга в различных ситуациях с помощью разнообразных жестов, поз, мимики.
5.4.

