 2015-05-06
2015-05-06 376
376Мы подошли вплотную к вопросу о том, каким образом в эволюции наследственно фиксируются новые частные реакционные механизмы приспособительного значения, т. е. каким образом внешний фактор развития замещается внутренним, т. е. реакционный механизм (ответ на внешнее раздражение) замещается корреляционным механизмом (ответ на внутреннее раздражение).
На примерах выпадения излишних реакций мы рассмотрели явление лишь кажущегося закрепления модификаций, получающееся в результате перехода организма в иную среду, в условиях которой эти реакции фактически не осуществлялись. Наличие нескольких наследственно обусловленных реакций на разные условия среды производит впечатление широкой изменчивости «под влиянием» внешних факторов, а если все реакции, кроме одной, оказываются утерянными, то возможность осуществления лишь одной определенной реакции производит впечатление большой наследственной устойчивости данной формы. В самом деле, здесь не было приобретения чего-либо нового, и кажущееся «фиксирование» есть лишь результат распада старых реакционных, а следовательно, и корреляционных систем, потерявших свое значение.
Однако совершенно ясно, что мы не можем объяснить эволюции организмов одним лишь распадом наследственных механизмов. Мы должны допустить, что, наряду с распадом реакционных и корреляционных систем, потерявших свое значение в новой среде, происходит и созидание новых наследственных механизмов. В предыдущей главе мы ознакомились с явлениями приспособления организмов к внешней среде и убедились, что в этом процессе приспособления, кроме естественного отбора более «пассивных» признаков, не имеющих существенного значения в прогрессивной
Глава IV. Прогрессивная эволюция. Адаптациогенез 161
эволюции, происходит приобретение новых реакционных механизмов общего значения. Вместе с тем происходит и дальнейшая пифференцировка таких механизмов общего значения (например, общая реакция опорных тканей на действие мышечной) на все более частные, локальные (например, усиленная работа определенных мышц вызывает разрастание определенных частей скелета). Эта дифференциация представляет в основном процесс непосредственного приспособления, т. е. имеет характер модифика-ционных изменений.
Однако на общем фоне этих модификациоиных изменений, несомненно, очень постепенно вырабатываются новые наследственные механизмы, определяемые как внешними, так и внутренними факторами развития, или, как мы их называли,— новые реакционные и корреляционные системы. Особое значение имеет для нас как раз процесс исторической замены внешних факторов развития внутренними, так как эта замена приводит к «автономности» развития нового признака, к известной независимости его от случайных колебаний факторов внешней среды, т. е. к максимальной его стабильности. В этом случае и принято говорить о прочном наследовании ('признака».
Мы уже обратили внимание на тот факт, что при возвращении исторически модифицированного организма в условия его исходной среды он обычно лишь частично приобретает прежние формы. Модификационное изменение оказывается в более или менее значительной мере стабилизированным («наследственным») и осуществляется в значительной мере независимо от тех факторов, которые первоначально его вызвали.
Факты стабилизации такого рода широко известны. Ламаркисты постоянно (начиная с Нэгели) ссылались на наблюдаемый «параллелизм» наследственных изменений природных экологических или географических форм и модификациоиных изменений близких форм, в условиях эксперимента, как на доказательство прямого унаследования результатов индивидуального приспособления. Этот вопрос обсуждался у нас в Союзе П. В. Серебров-ским [1928] с ламаркистских позиций, М. А. Розановой [1926], Е. И. Лукиным [1936, 1940] —с дарвинистических. У меня факты такого рода сопоставлены в виде таблички [И. И. Шмальгау-зен, 1939—1940]. Приведем некоторые более новые примеры.
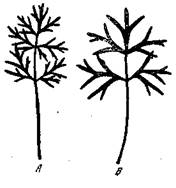
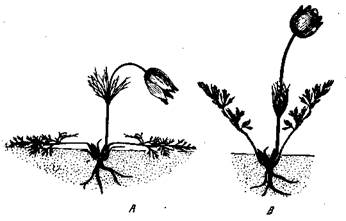
Циммерман [Zimmermann, 1938] получал у обыкновенной Anemone Pulsatilla путем черенкования от одной и той же особи в разных условиях освещения две модификации — теневую, с тонко рассеченными листьями, и световую (на солнце) — с мало рассеченными листьями (рис. 25). Этим модификациям соответствуют наследственные расы той же Anemone Pulsatilla — западная раса с тонко рассеченными листьями и восточная —с мало рассеченными листьями, которые при совместном воспитании в одних и тех же условиях влажности и освещения дают резко
v И. и. Щмальгаузец
162 Организм как целое в индивидуальном и историческом развитии
различные формы (рис. 26). Эти изменения имеют приспособительный характер, определяя оптимальную транспирацию растения в разных условиях влажности (что было опять-таки проверено путем эксперимента).
Н. И. Калабухов и В. М. Родионов [1938] показали, что равнинные предкавказские лесные мыши при перенесении в горы
|
Рис. 25. Модификации формы листа у анемоны (сон — Anemone Pulsatilla) [Циммерманн, 1938]
А — теневая модификация; В — световая модификация
Рис. 26. Географические формы анемоны (Anemone Pulsatilla) [Циммерманн, 1938]
А — западная форма, ssp. germanica; В — восточная форма, ssp. grandis
дают типичную горную реакцию в виде увеличения числа эритроцитов и количества гемоглобина в крови. Однако горные формы того же подвида: Apodemus sylvaticus ciscaucasicus, произошедшие очевидно, от равнинной мыши в результате такой индивидуальной адаптации, при перенесении на низменность уже не дают обратной реакции. Повышенное содержание гемоглобина у них наследственно стабилизировалось.
Процесс постепенной стабилизации функциональных признаков организации и развития в процессе эволюции автономного аппарата онтогенеза показан А. А. Машковцевым на сравнительно эмбриологическом материале по развитию ряда органов у но-
/ лава Прогрессивная Эволюция. Адаптацидгёнёз 163
звоночных [1936]. Следуя В. Ру, А. Машковцев различает до-функциональный период самодифференцирования органа и функциональный период, когда дифференцирование является процессом, зависимым от функции. Дофункциональный период дальше подразделяется на ряд типичных этапов развития: этап лабильной детерминации, этап самодифференцировки и этап зависимого развития (от гуморальных факторов). Существование таких этапов показано экспериментально на ряде органов (главным образом органы выделения и органы дыхания), и затем проведено сравнение относительного значения этих этапов в морфогенезе различных позвоночных. Оказалось, что можно установить совершенно закономерные сдвиги значения факторов онтогенеза в филогенетическом ряду позвоночных. И в развитии, по крайней мере вторичных, почечных канальцев и мочеточника, и в развитии легочных альвеол первоначально, у низших позвоночных, решающую роль играет сама функция, без которой эти структуры не развиваются. При переходе к филогенетически более дифференцированным формам формообразовательное значение функции падает и сдвигается на более поздние стадии. У аксолотля фрагментация легкого и образование альвеол идет исключительно под влиянием функции, у лягушек в дофункциональный период намечается лишь первичная фрагментация легкого. У жаб, как животных, еще более приспособленных к наземной жизни, альвеолярная структура легкого развивается еще в дофункциональный период, именно у личинки, которая у жаб легкими совершенно не дышит. В функциональный период происходит дальнейшее усложнение строения легких. Первоначальная фрагментация легкого происходит у жабы не под влиянием функции, но под влиянием гормона щитовидной железы, который определяет целый ряд морфогенетических процессов во время метаморфоза амфибий. У рептилий и у млекопитающих наблюдается типичное «самодифференцирование» легочных структур в эмбриональный период. Функция начинается, естественно, только после рождения и ведет к дальнейшему новообразованию структур. Только клоачные и сумчатые млекопитающие составляют исключение в том смысле, что у них функция сохранила еще значение основного формообразовательного, фактора, и альвеолы легкого развиваются только под влиянием начинающегося дыхания.
Таким образом, ясно видно формообразовательное значение функции у филогенетически молодых органов, затем значение формообразовательного фактора иногда переходит к эндокринным, т. е. во всяком случае внутренним факторам развития организма, и, наконец, зависимое развитие может более или менее уступить место «самодифференцированию», т. е. «автономному» развитию, которое зависит в основном от внутренних факторов, заложенных в самой ткани данного зачатка (по крайней мере после периода его полной детерминации). Картина постепенной
1б4 Организм как целое в индивидуальном и историческом развитии
замены внешних факторов развития внутренними и затем переноса этих внутренних факторов внутрь самого зачатка довольно ясна. Конечно, структура легких, даже у примитивных амфибий, не возникает под влиянием простого растяжения, хотя картина фрагментации легкого у низших амфибий весьма напоминает такое растяжение стенок между развивающейся сетью кровеносных сосудов, являющейся как бы механическим препятствием. По сосудам идет затем развитие легочных трабекул. Однако так примитивно мы себе не будем этого представлять на данном этапе развития органа. Ведь это далеко не первое возникновение нового органа. Легкое амфибий имеет за собой уже длительную историю последовательных фаз прогрессивной дифференцировки. Здесь мы имеем, конечно, уже наследственно «зафиксированную» структуру. Однако внешний фактор — функция (и хотя бы даже механическое растяжение легочного мешка) все еще является в роли освобождающего раздражителя. Нужно думать, что раньше определяющая роль функции была еще выше. Во всяком случае указанный ряд дает нам вполне конкретную картину смены внешних факторов развития внутренними.
На те же явления стабилизации указывают и вообще факты автономного развития многих функциональных структур (структуры кости, формы мускульных гребней и отростков костей, соединительнотканных структур), которые филогенетически развивались в значительной мере под непосредственным влиянием функции. Так, например, специфические функциональные структуры механического слоя кожного покрова взрослого животного закладываются на ранних стадиях эмбрионального развития, как уже детерминированные внутренними факторами, независимо от местных механических влияний, которые не сравнимы с механической нагрузкой кожи в различных частях тела взрослого животного [А. Браун, 1939].
Начальной фазой автономизации развития является авторегу-ляторное развитие, допускающее известный диапазон колебаний интенсивности внешнего фактора для наступления нормальной реакции адаптивного характера [Шмалъгаузен, 1940]. Нельзя не указать при этом, что даже в том случае, когда внешние факторы, с которыми связано развитие известного признака, полностью сохраняют еще свое детерминирующее значение, оно может быть ограничено очень коротким временем, после которого индивидуальное развитие идет в значительной мере автономно (в отношении данного признака). Таково детерминирующее значение низких температур для вегетации растений средних широт (на стадии яровизации) и детерминирующее значение освещения (на световой стадии). В самом существовании стадии яровизации нельзя не видеть особой формы защиты растения от «ошибочного» прорастания в условиях теплой осени [Лысенко, 1936]. Световая и теневая форма листьев индуцируется на деревьях иногда
Глава IV. Прогрессивная эволюция. Адаптациогбнёз 165
еще в почках, заложившихся в предыдущем году, в зависимости от условий более интенсивного летнего освещения, а не слабого весеннего, когда почка развертывается [Nordhausen, цит. по: lingerer, 1926]. И в этом мы видим «защиту» организма от «ошибочных» реакций на переменчивые условия весеннего освещения, т. е. известные черты автономизации развития, которые объяснимы только естественным отбором, т. е. элиминацией всех неблагоприятных уклонений (т. е. «ошибочных» модификаций).
Более полная автономизация развития определяется усложнением системы коррелятивных зависимостей между онтогенетическими процессами, т. е. усложнением аппарата индивидуального развития. В особенности большое значение имеет создание регу-ляторного аппарата, «защищающего» нормальное формообразование от его нарушений при нередких уклонениях в факторах внешней среды. Этот регуляторный аппарат достигает, как мы уже видели, особенно большой сложности у позвоночных животных (система градиентов, индукционные системы, морфогенетические и гуморальные взаимозависимости и, наконец, эргонтические корреляции) и допускает существование довольно значительных сдвигов как в интеысивностях факторов внешней среды, так отчасти и во внутренних факторах развития.
Наконец, и сама доминантность нормальных признаков есть результат развития морфогенетического аппарата, обладающего двойным «запасом» активности, обеспечивающим развитие нормального фенотипа и у гетерозиготы (т. е. в половинной «дозе»). Эта точка зрения была развита рядом генетиков, в особенности Мёллером, и разработана М. Камшиловым [1940] в новом аспекте. Доминирование нормы и рецессивность мутаций мы рассматриваем как одно из первых выражений стабилизации форм.
Все описанные регуляционные механизмы делают организм более устойчивым, более стабильным по отношению к факторам, которые могли бы повлиять на процессы онтогенеза. Мы объясняем стабилизацию форм действием естественного отбора, неразрывно связанного с постоянной элиминацией всех неблагоприятных уклонений от приспособленного «нормального» фенотипа, каковы бы ни были источники этих уклонений. Элиминация всех «ошибочных» реакций на временные уклонения в факторах среды ведет к выработке более автономного аппарата индивидуального развития, т. е. к стабилизации форм.
Мы привели много примеров, указывающих на ведущее значение модификационных изменений, которые в процессе эволюции затем стабилизируются. Что эта стабилизация не есть результат простого наследственного «закрепления» реакции в ряду поколений, т. е. что эти факты не поддаются ламаркистскому истолкованию, видно из следующего. Многие наземные растения выдерживают более или менее длительное содержание под водой. При этом наблюдаются многие адаптивные изменения, больший-
166 Органам как целое в индивидуальном и историческом развитии
стВо из которых можно рассматривать как явления недоразвития или редукции [см. Ungerer, 1926]. У амфибиотических растений такие реакции принимают более строго специфический характер. Адаптивная модификация возникла, бесспорно, как приспособление к водной среде, и если бы она прямо фиксировалась как таковая, то соответствующая морфогенетическая реакция должна была бы определяться именно развитием в водной среде. Однако у водяной гречихи (Polygonum amphibium) можно получить типичные «плавающие» листья во влажном воздухе, а не только в воде. У стрелолиста (Sagittaria sagittifolia) типичные лентовидные водяные листья развиваются и в воздухе при ослабленном освещении [К. Goebel, 1928]. Таким образом, детерминирующим фактором развития здесь является не тот фактор среды, который вызвал к жизни данную модификацию. В процессе эволюции выработался новый механизм индивидуального развития, что, очевидно, возможно только через естественный отбор. Так как отбор происходил на фоне установившейся адаптивной модификации, то он, во всяком случае частично, проявлялся в форме стабилизирующего отбора. В данных случаях известная степень влажности и затененности были, очевидно, более постоянными и более надежными раздражителями, чем переменчивый уровень воды в заболоченной местности. Смена детерминирующих факторов произошла в результате элиминации всех неудачных реакций (т. е. ошибочных адаптации).
Подобная смена формативных раздражителей известна и для ксерофитов. Если у них имеются два рода листьев: первичные — теневые и вторичные — ксерофитные, то первичные развиваются при преобладании минерального питания, а вторичные при преобладании ассимиляции. Получается парадоксальное явление — ксерофитные листья развиваются не только при образовании корней или при сильном освещении, но и при культуре в чистой воде [W. Vischer, цит, по: Goebel, 1928]. И здесь ксерофитный характер листьев, возникший как модификация, приспособленная к засушливым условиям, развивается затем под влиянием иных внешних факторов, обеспечивающих, очевидно, более надежное наступление формообразовательной реакции (независимо от случайных колебаний влажности). И здесь имела место настоящая наследственная стабилизация через отбор, а не простое фиксирование механизма адаптивной модификации. Естественный отбор в этих случаях был связан с постоянной элиминацией всех неадекватных форм реагирования и, в частности, всех «ошибочных» модификаций.
Глава IV. Прогрессивная эволюция. Адаптациогенеэ 167








