2015-05-06
2015-05-06 452
452Такая регуляция обычно наблюдается очень ясно на ранних стадиях развития зародыша, но в известной степени даже у взрослых организмов (в особенности у растений и низших животных). У растений, многих кишечнополостных животных (гидроиды и медузы), иглокожих (морские звезды и ежи), ланцетника и у всех позвоночных животных можно отделить друг от Друга первые клетки зародыша, и каждая из этих клеток может Дать начало нормально сформированному (только уменьшенному) зародышу и затем развиться в совершенно нормальное взрослое животное или растение. У гидромедуз один из восьми первых бластомеров может дать начало миниатюрной, но вполне нормальной личинке. Даже части зародыша на стадии бластулы могут еще развиться в целые личинки. И у человека каждый из первых двух бластомеров может развиться в нормального зародыша. Таким образом возникают нередко наблюдаемые «однояйцовые близнецы», которые поражают своим сходством.
Регуляционные яйца имеют более или менее однородную цитоплазму, поэтому продукты их деления сходны и содержат все материалы целого яйца. Однако, как бы ни была совершен-
240 Регуляция формообразования в индивидуальном развитии
на способность частей таких регуляционных яиц к восстановлению целого, она не безгранична. У животных яйца всегда имеют некоторую, по меньшей мере полярную организацию. Субстанции цитоплазмы, хотя и не строго локализованы, но имеют разную концентрацию на разных уровнях яйца, это определяет некоторые различия в дальнейшем развитии анимальной и вегетативной его половин. Полярная организация яйца связа-
|
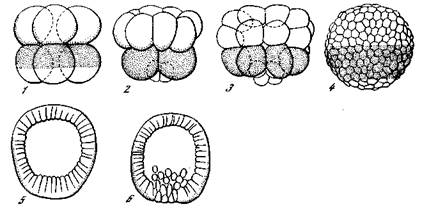
| Рис. 4. Дробление (1 — 5) и начало образования первичной мезенхимы (6) у морского ежа Paracentrotus lividus [Кюн, 1955] |
на с положением яиц в составе зачаткового эпителия яичника. Она отличается поступлением запаса питательного материала в одном определенном направлении, которое становится главной осью яйца. Наиболее обстоятельно изучены явления регуляции на ранних стадиях развития у морских ежей (рис. 4) и земноводных позвоночных (см. главу II).
Яйцо морского ежа (Paracentrotus lividus) имеет шарообразную форму. Под экватором, т. е. в вегетативной половине яйца, располагается пигментированное кольцо, отмечающее область будущей энтодермы, а под ним, в районе вегетативного полюса,— сегмент прозрачной цитоплазмы, отмечающий область будущей мезенхимы (клеток — образователей скелета). Яйцо имеет, следовательно, радиально-симметричную организацию. Так как первые две плоскости деления дробления проходят вертикально, то первые четыре бластомера, получающиеся в результате этих делений, совершенно равнозначны. Они содержат все материалы яйца в тех же соотношениях, какие были и в целом нераздробленном яйце. При обособлении бластомеров меняются, однако, их соотношения с внешней средой. В норме первые два бластомера тесно соприкасаются своими внутренними стенками, развившимися в результате клеточного деления. Это вызывает различия в строении наружных слоев протоплазмы. Наружная я внутренняя стенки попадают в разное положение по отношению
Глава 1. Детерминированное развитие и регуляция 241
к процессам обмена (например, поглощению кислорода и выделению некоторых продуктов обмена), которые возможны только через внешнюю поверхность. Очевидно, с этим связано и направление дальнейшей их дифференцировки. Каждый бластомер в норме дает начало только половине зародыша. При искусственном отделении бластомеров они округляются и принимают вновь ту же радиально-симметричную структуру, которую имело и не-раздробившееся яйцо. То же самое происходит и при разделении первых четырех бластомеров. Поэтому каждый из этих четырех бластомеров при их изоляции дает начало совершенно нормальной (но уменьшенной) личинке. После двух вертикальных делений яйца третья плоскость деления проходит горизонтально и отделяет верхнюю — анимальную — половину яйца от нижней — вегетативной. Продукты деления оказываются уже различными. Четвертое деление протекает вновь в вертикальной плоскости и приводит к образованию в анимальной половине 8 одинаковых клеток, а в вегетативной — 4 крупных пигментированных клеток (макромеров), включающих основную массу желтка, и 4 небольших клеток (микромеров) с прозрачной цитоплазмой. При отделении анимальной половины яйца из нее развивается только эктодермальный пузырь, а затем развитие останавливается. Из вегетативной половины может развиться почти нормальная личинка. Если удалить только небольшие микромеры вегетативного полюса, то личинка не развивается. С другой стороны, стоит прибавить к клеткам анимальной половины хотя бы два микромера, как из этих клеток развивается нормальная личинка.
Серией последовательных опытов комбинирования бластомеров, а также опытами перемещения субстанций цитоплазмы яйца (центрифугированием) доказано, что пигментация вегетативной половины не имеет никакого формообразовательного значения, а ее желток является только питательным материалом. Однако прозрачная цитоплазма вегетативного полюса, входящая в состав упомянутых микромеров, содержит какие-то весьма активные вещества, влияющие на развитие всего зародыша. Эти вещества содержатся внутри клеток и определяют развитие не только тех клеток, в которых они находятся (микромеры и развивающаяся из них мезенхима), но оказывают формообразующее воздействие и на другие части зародыша. В этом и заключается основное различие между мозаичными и регуляционными яйцами.
Во всех типах яиц имеется своя унаследованная организация а имеются различные необходимые для развития субстанции, которые распределяются при клеточных делениях и определяют в известной мере дальнейшую судьбу тех клеток, в которые они попадают. Различие состоит, однако, в том, что одни такие вещества удерживаются внутри клеток и влияют только на диф-ференцировку этих клеток, а другие вещества (или продукты
242 Регуляция формообразования в индивидуальном развитии
обмена, выделяемые в результате их использования) проникают через клеточную оболочку в межклеточные пространства и в соседние клетки и ткани. Взаимодействие частей, сопровождающееся передачей активных веществ от одной части зародыша к другой, и составляет главную особенность регуляционного развития (в узком смысле термина регуляции).
Регуляция наблюдается не только на стадиях дробления яйца. Ранние зачатки целых органов могут быть искусственно расчленены, и из отдельной части в подходящих условиях могут развиться вполне нормальные органы (например, зачатки глаз или конечностей). Необходимо, однако, сохранение определенных связей (например, эпителия и мезенхимы в зачатке конечностей), которые имеют решающее значение в развитии данного органа. В системе развивающегося органа регулируется и соотносительный рост, и уровень дифференцировки частей (например, соотношение между глазным бокалом и хрусталиком глаза), как это доказано многими опытами искусственного вмешательства — удаления частей, пересадок и т. п. Многие такие регуляции возможны и у взрослых особей. Примером может служить восстановление целого органа из небольшой его части (регенерация конечностей у земноводных, хвоста у ящериц и т. п.), целого организма из отдельной части (у гидроидов, планарий, ас-цидий) или даже целого организма из отдельной клетки (у растений — например, из клетки листа бегонии).
Во всех этих случаях решающее значение имеют нарушение и восстановление некоторых связей, которые определяют развитие взаимодействующих частей. Взаимовлияние частей может выражаться в возбуждении одних формообразовательных процессов и в торможении других. Вопросам о природе этих влияний мы и посвятим дальнейшее изложение.
Глава II
РЕГУЛЯЦИЯ ФОРМООБРАЗОВАНИЯ У ЗЕМНОВОДНЫХ
Яйца земноводных (лягушки, тритоны, аксолотль) очень удобны для исследования. Они достаточно велики, легко освобоЖ-даются из оболочек и весьма выносливы при экспериментировании. Молодые зародыши допускают разделение на части, их перекомбинирование, пересадку зачатков в другое место того же или другого зародыша. Можно производить и обмен зачатками между зародышами разных видов, отличающихся по величй-
Глава II. Регуляция формообразования у земноводных 243
ре или пигментации клеток. Большое значение имеет также возможность культивирования частей зародыша, отдельных зачатков и их комбинаций в солевых растворах. Все это возможно, так как клетки молодых зародышей амфибий достаточно устойчи-вЫ и снабжены своим собственным питательным материалом в виде рассеянных в их цитоплазме желточных зерен. Поэтому у земноводных, формообразовательные процессы изучены лучше, чем У каких-либо других животных. Результаты этого исследования имеют общее значение. Это ясно показывают значительно более трудные, но все же достаточно многочисленные эксперименты с яйцами и зародышами различных рыб, птиц и даже млекопитающих.
Зрелое яйцо амфибий имеет заложенную еще в яичнике полярную организацию и некоторые признаки двусторонней симметрии. Это выражается в более значительной концентрации крупных желточных зерен в вегетативной половине и меньшей концентрации очень мелких желточных зерен у анимального полюса. В переходной зоне, в районе экватора, и в центральной массе протоплазмы преобладают мелкие и средние желточные зерна. В поверхностном слое анимальной половины яйца находится темный, иногда совсем черный пигмент. Кроме желточных зерен, цитоплазма содержит многочисленные зернистые структуры (митохондрии, липохондрии) и химические вещества (рибонуклеотиды, фосфатиды и др.).
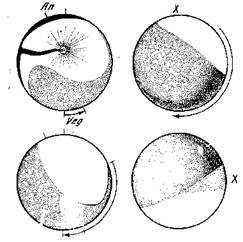
Двусторонняя симметрия намечается вначале только некоторым распространением пигментированного материала в направлении будущей брюшной стороны зародыша. Окончательная структура яйца в норме устанавливается после проникновения сперматозоида. В экспериментальных условиях она еще может быть изменена. После оплодотворения сейчас же возникают токи цитоплазмы, перемещающие желточные массы вегетативного полюса. Так как желток имеет более значительный удельный вес, то и все яйцо несколько поворачивается. Одновременно происходит смещение поверхностного слоя цитоплазмы. В результате этого между пигментированной и непигментированной поверхностью яйца в районе экватора образуется серповидная промежуточная зона, получившая название серого полумесяца (рис. 5). Эта зона отмечает область будущей спинной стороны зародыша.
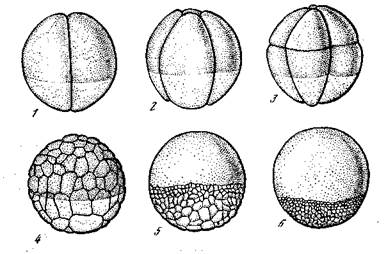
При дроблении яйца (рис. 6) первые плоскости дробления Проходят вертикально, однако без определенной связи с имеющейся уже плоскостью симметрии. Последующими делениями яйцр дробится синхронно на 4, 8, 16 клеток. Вегетативные клетки крупнее анимальных, и их дальнейшие деления запаздывают По сравнению с анимальными. Разница в величине клеток поэтому возрастает. Образуется стадия полого пузырька — бластула с толстой, крупноклеточной нижней стенкой и более тонкой, мел-коклеточной, верхней. Вслед за сформированием бластулы раз-
244 Регуляция формообразования в индивидуальном развитии
растается анимальная ее половина, которая по экватору надвигается на вентральную. Вслед за тем в районе серого полумесяца начинаются перемещения клеток, ведущие к образованию щелевидного углубления (бластопора), через верхний край которого клетки анимальной половины вворачиваются внутрь. Клетки анимальной половины все более распространяются по поверхности яйца и охватывают вегетативную половину. Наиболее энергичные движения происходят в спинной области, между анимальным полюсом и упомянутым краем бластопора. Эти движения сливаются в поток клеток, направляющихся назад, к верхнему краю — губе бластопора. Здесь этот поток клеток заворачивает через край бластопора внутрь и продолжает свое
|
Рис. 5. Образование серого полумесяца в яйце лягушки [Кюн, 1955]
Стрелки — поворот яйца после проникновения сперматозоида; An — анимальный полюс; Veg — вегетативный полюс; X — серый полумесяц
Рис. 6. Дробление яйца Rana silvatica [Кюн, 1955]
1 —з — 2-, 4- и 8-клеточная стадия; 4 — крупноклеточная бластула; 5, 6 — мелкоклеточная бластула
|
| Глава II. Регуляция формообразования у земноводных 2'г |
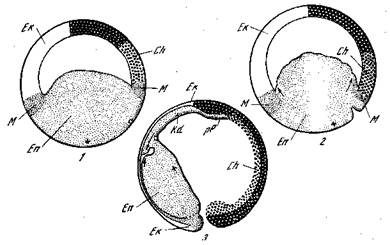
Рис. 7. Гаструляция у тритона Pleurodeles waltlii [Кюн, 1955]
1 — бластула; 2 — ранняя гаструла; 3 — ранняя нейрула; Ch — хорда; Ek — экто
дерма; En — энтодерма; Кб, — головная кишка; М — мезодерма; рР — прехордаль-
ная пластинка; Н---- место вегетативного полюса
движение в обратном направлении — вперед, образуя непрерывный второй слой клеток, подстилающих наружный слой клеток спинной области. Край бластопора, имеющий вначале вид небольшой поперечной щели, расширяется по краю клеток ани-мальной половины: он приобретает форму широкой дуги, которая по мере обрастания клеток вегетативной половины превращается в кольцо, все более стягивающееся над вегетативными клетками, которые таким образом попадают внутрь зародыша. По всему этому краевому кольцу происходит, однако, не просто надвигание анимальных клеток, но и их перемещение внутрь, т. е. тот же процесс, который начался в области серого полумесяца. Крупные вегетативные клетки при всех этих движениях играют довольно пассивную роль.
В результате этих первых формообразовательных движений однослойный полый пузырек — бластула — превращается в гаст-РУлу (рис. 7), т. е. в двухслойный зародыш, имеющий уже первичную кишку (энтодерму) со своей полостью и первичные покровы (эктодерму). Нижняя, вентральная стенка первичной кишки составляется из крупных набитых желтком клеток вегетативной половины яйца (собственно энтодерма). Верхняя, дорсальная, стенка, так называемая крыша первичной кишки, составляется из клеток того потока, который ввернулся со сцйнной поверхности анимальной половины внутрь через верх-
246 Регуляция формообразования в индивидуальном развитии
ний край бластопора (это зачаток будущей хорды и мезодермы). Эти формообразовательные движения клеток предопределены структурой плазматического содержимого яйца.
Крупные желточные зерна вегетативного полушария яйца являются запасом питательного материала, который позднее входит в состав стенки кишечника и обеспечивает развитие зародыша вплоть до его вылупления из оболочек и формирования личинки, способной к самостоятельному питанию. В ани-мальном полушарии содержится гораздо меньшее количество и притом очень мелких желточных зерен. Кроме того, цитоплазма анимальной половины яйца содержит значительно больше активных веществ — энзимов и рибонуклеиновых кислот, нуклео-тидов и фосфатидов. Большое значение приписывается и белкам, содержащим сульфгидрильные группы (SH-группы), которые концентрируются на анимальном полюсе в районе, где происходят деления созревания яйца. После этого как сульфгидрильные белки, так и рибонуклеиновые кислоты концентрируются главным образом на спинной стороне анимального полушария.
Перемещения клеток в процессе гаструляции приводят к совершенно новым соотношениям. Вегетативные клетки, которые раньше непосредственно соприкасались с внешней средой, оказываются внутри зародыша, ограничивая просвет первичной кишки. Анимальные клетки распространяются по всей поверхности зародыша, образуя его первичные покровы (эктодерму). Однако анимальные клетки краевой зоны, и особенно спинной части оказываются теперь внутри зародыша, составляя крышу первичной кишки, или точнее зачаток хордомезодермы. Этот слой клеток вступает теперь в контакт с поверхностным слоем эктодермы. Раньше под слоем клеток анимального полюса находилась заполненная жидкостью полость бластулы. Теперь в спинной части зародыша под внешним слоем клеток располагается не полость, а второй слой клеток, соответствующий крыше первичной кишки (хордомезодерме), под средней частью которой возникает полость первичной кишки — гастроцель.
Изменение связи с внешней средой (поступление кислорода!) должно привести к изменению метаболизма поверхностных клеток (эктодермы), с одной стороны, и лежащих теперь в глубине, но все же активных клеток крыши первичной кишки (хордомезодермы) — с другой. Естественно, что между этими двумя клеточными пластами должен происходить обмен, по меньшей мере передача кислорода от поверхностных клеток эктодермы к внутренним слоям и передача некоторых веществ от клеток хордомезодермы к клеткам эктодермы.
Таким образом, неизбежно должно установиться физиологическое взаимодействие между слоями клеток, различающихся по своему метаболизму. Это взаимодействие может, очевидно, оказать влияние и на дифференцировку клеток и на формообразо-
Глава II. Регуляция формообразования у земноводных 247
вание соответствующих частей тела. В действительности, начиная с установления контакта между эктодермой спинной области зародыша и крышей первичной кишки, судьба этих частей оказывается предопределенной. Из крыши первичной кишки развивается по средней линии спины центральная скелетная ось — хорда (вокруг которой позднее разовьется позвоночник и задняя часть черепа), а по бокам от нее — мезодерма (из которой позднее разовьются сегментированные боковые мышцы, стенки общей полости тела, почки, половые железы и др.).
Из дорсальной эктодермы, прилегающей к крыше первичной кишки, развивается утолщенная полоса эктодермальных клеток — нервная пластинка (т. е. общий зачаток нервной системы). Образование нервной пластинки является также результатом активного перемещения клеток: клетки нижнего слоя эктодермы анимальной половины гаструлы стягиваются по направлению к средней линии спинной стороны зародыша.
Можно было бы предположить, что здесь опять перед нами внутриклеточное влияние формообразовательных веществ яйца, которые лишь перераспределились при описанных перемещениях клеточного материала. Многочисленные эксперименты показывают, однако, что это не так. Если перемещение материала и было уже предопределено организацией яйца, то новые его дифферен-цировки определяются именно установившимися новыми соотношениями и, в первую очередь, взаимодействием между эктодермой и хордомезодермой.